
Abstrak
- Di daerah beriklim sedang, musim dingin bisa menjadi masa yang sulit bagi serangga eusosial. Kelangsungan hidup koloni lebah madu selama musim dingin bergantung pada keseimbangan yang baik antara pengaturan suhu sarang, pengelolaan cadangan makanan, dan waktu dimulainya generasi lebah pekerja baru. Kelangsungan hidup di musim dingin dipengaruhi oleh beberapa faktor, terutama ukuran koloni, tingkat serangan tungau Varroa, dan ketersediaan makanan yang disimpan. Yang terpenting, kondisi iklim dan sumber daya bunga pada musim mencari makan sebelumnya juga dapat memengaruhi kesehatan lebah madu dan kekuatan koloni sebelum hibernasi.
- Studi ini, yang dilakukan di seluruh Eropa, meneliti bagaimana komposisi lanskap dan kondisi cuaca memengaruhi kelangsungan hidup koloni lebah madu di musim dingin. Studi ini menggunakan keragaman serbuk sari sebagai proksi untuk kualitas sumber daya bunga dan hari-hari mencari makan yang tersedia sebagai variabel iklim untuk memahami hubungan kausalnya dengan kelangsungan hidup di musim dingin.
- Kami menemukan bahwa lanskap dengan persentase area pertanian yang lebih tinggi meningkatkan keanekaragaman serbuk sari yang dikumpulkan oleh lebah madu di musim gugur, sedangkan persentase area semi-alami yang lebih tinggi meningkatkan keanekaragaman selama musim panas. Keanekaragaman serbuk sari di musim semi dan musim gugur merupakan pendorong utama keberhasilan bertahan hidup di musim dingin, yang menekankan pentingnya sumber daya bunga yang beragam bagi kesehatan koloni. Meskipun kami tidak menemukan pengaruh cuaca yang signifikan secara statistik terhadap kelangsungan hidup di musim dingin, tren menunjukkan adanya pengaruh potensial, yang menjamin penelitian lebih lanjut untuk mengonfirmasi dan mengklarifikasi peran pencarian makanan musiman terhadap kesehatan koloni.
- Sintesis dan aplikasi : Studi kami menyoroti peran penting dari penyertaan keragaman sumber daya bunga dan kondisi cuaca dalam kerangka kerja komprehensif untuk mempelajari hibernasi lebah madu. Studi ini menunjukkan bahwa peningkatan keragaman tanaman di sekitar tempat pemeliharaan lebah dan penerapan praktik pertanian yang meningkatkan sumber daya bunga dapat secara signifikan meningkatkan kelangsungan hidup di musim dingin, dengan koloni lebah madu yang diuntungkan bahkan di lanskap dengan aktivitas pertanian yang lebih tinggi, berbeda dari kebutuhan penyerbuk lainnya.
1. PENDAHULUAN
Di daerah beriklim sedang, serangga eusosial abadi menghadapi tantangan untuk bertahan hidup dalam periode suhu dingin yang panjang dan sumber daya yang langka. Misalnya, strategi hibernasi lebah madu Barat, Apis mellifera , tidak hanya melibatkan mempertahankan suhu optimal di dalam sarang tetapi juga dengan hati-hati mengatur waktu penghentian dan dimulainya kembali pemeliharaan induk, bersama dengan pengelolaan persediaan makanan yang efisien (Seeley & Visscher, 1985 ). Akibatnya, perhatian dan penelitian yang ekstensif telah didedikasikan untuk menjelaskan kompleksitas lebah madu yang melewati musim dingin (Döke et al., 2015 ; Minaud et al., 2024 ) karena tingkat kelangsungan hidup dan kesehatan koloni di musim semi secara langsung memengaruhi produksi madu dan efektivitas dalam penyerbukan tanaman dan tumbuhan liar (Geslin et al., 2017 ).
Strategi hibernasi untuk bertahan hidup di musim dingin dimulai pada akhir musim panas, saat koloni telah menyimpan cukup persediaan musim dingin (nektar dan serbuk sari) dan ratu lebah madu mulai membesarkan lebah ‘musim dingin’, yang berbeda dari lebah musim panas baik secara fisiologis (Fluri et al., 1977 ; Hatjina et al., 2014 ) dan dalam rentang hidup (Mattila et al., 2001 ). Karena suhu secara konsisten turun di bawah 15 derajat Celsius, pemeliharaan induk berhenti dan koloni membentuk klaster yang akan mampu mempertahankan suhu dan kondisi kelembapan yang stabil di dalam sarang sepanjang musim dingin (Fahrenholz et al., 1989 ; Stabentheiner et al., 2003 ). Sementara strategi bertahan hidup musim dingin yang berhasil melibatkan pembentukan klaster dan mempertahankan kondisi yang stabil di dalam sarang, beberapa faktor dapat menyebabkan kegagalan koloni musim dingin. Hal ini termasuk ukuran koloni yang tidak mencukupi saat memasuki musim dingin (Döke et al., 2019 ), koloni yang lebih kecil kurang efektif dalam pengaturan suhu tubuh (Free & Racey, 1968 ), serangan hama Varroa destructor yang tidak diobati sehingga melemahkan koloni sebelum musim dingin (van Dooremalen et al., 2012 ), dan kelaparan karena cadangan makanan yang tidak memadai (Stabentheiner et al., 2003 ). Stresor tersebut, baik secara individu maupun kombinasi, pada akhirnya dapat membahayakan kemampuan koloni untuk bertahan hidup di bulan-bulan musim dingin (Döke et al., 2015 ).
Faktor-faktor yang disebutkan di atas merupakan konsekuensi dari kondisi lingkungan dan kondisi kesehatan koloni pada musim tanam sebelumnya, sehingga sulit untuk mempelajari kelangsungan hidup di musim dingin sebagai hasil terpisah dari beberapa parameter yang diukur tepat sebelum pembentukan klaster. Oleh karena itu, perlu untuk mempertimbangkan struktur kausal yang lebih luas untuk memperoleh pemahaman yang komprehensif tentang kelangsungan hidup di musim dingin.
Ketersediaan sumber daya bunga sangat penting bagi koloni lebah madu. Penyederhanaan lanskap, yang didorong oleh berbagai faktor, seperti intensifikasi pertanian dan urbanisasi (Goulson et al., 2015 ), dapat mengurangi kekayaan dan kompleksitas komposisi komunitas tanaman (Power et al., 2012 ). Pada gilirannya, hilangnya sumber daya bunga dan habitat yang sesuai telah menjadi faktor yang berkontribusi terhadap penurunan penyerbuk (Brown & Paxton, 2009 ). Sumber daya bunga tidak hanya memengaruhi ketersediaan dan penyimpanan makanan, tetapi juga status kesehatan umum lebah individu karena kandungan nutrisi nektar dan, yang lebih penting, serbuk sari yang ditemukan pada spesies tanaman yang berbeda (Haydak, 1970 ; Roulston & Cane, 2000 ). Dalam koloni lebah madu yang dikelola, diet polifloral dan pengayaan lanskap sekitar dengan tanaman tangkapan melliferous ditemukan berkorelasi positif dengan peningkatan umum dalam fungsi imun (Alaux et al., 2010 ), toleransi terhadap penyakit (Pasquale et al., 2013 ) dan keadaan fisiologis lebah (Alaux et al., 2017 ). Namun, sedikit yang diketahui tentang efek komposisi lanskap pada kinerja koloni selama fase hibernasi koloni. Rutschmann et al. ( 2022 ) menemukan bahwa kelangsungan hidup koloni lebah madu liar selama musim dingin berhubungan positif dengan jumlah habitat semi-alami di sekitar lokasi sarang, yang juga menunjukkan bahwa efek lanskap dipengaruhi secara positif oleh sumber daya bunga. Dalam koloni yang dikelola, Requier et al. ( 2017 ) menemukan bahwa ketersediaan sumber daya serbuk sari di musim semi memiliki hubungan kausal tidak langsung dengan kelangsungan hidup koloni musim dingin melalui efek berjenjang di antara ciri-ciri demografi koloni. Meskipun demikian, dampak langsung dari keragaman bunga, yang diukur sebagai keragaman serbuk sari yang dikumpulkan oleh lebah madu pengumpul makanan, terhadap kelangsungan hidup koloni di musim dingin masih belum diketahui. Sama seperti sumber daya bunga, cuaca memainkan peran penting dalam perkembangan koloni dan dapat dikaitkan dengan kegagalan melewati musim dingin. Kondisi cuaca dari musim semi hingga musim gugur dapat secara langsung memengaruhi ukuran dan kesehatan koloni, karena lebah madu memerlukan kondisi yang baik untuk terbang mencari makan (Hatjina et al., 2014 ). Misalnya, peristiwa curah hujan ekstrem di musim semi dikaitkan dengan risiko yang lebih besar terhadap hilangnya populasi sebelum musim dingin dan berdampak negatif pada produksi madu (Quinlan et al., 2023 ). Mengenai kelangsungan hidup di musim dingin, pendekatan pemodelan matematika telah diadopsi untuk menggunakan data survei dari peternak lebah selama musim dingin berturut-turut dan untuk menentukan dampak dari beberapa faktor pendorong secara bersamaan. Musim panas yang lebih hangat dan lebih kering ditemukan sebagai prediktor signifikan kegagalan musim dingin (Calovi et al., 2021) .; Switanek et al., 2017 ). Demikian pula, musim gugur dan musim dingin yang lebih hangat diduga meningkatkan kemungkinan kegagalan karena seringnya terbang mencari makan dapat mengubah struktur klaster (Rajagopalan et al., 2024 ). Akan tetapi, studi eksperimental yang secara bersamaan menilai kondisi cuaca dan keanekaragaman serbuk sari sebagian besar belum ada.
Studi ini membahas kesenjangan ini dengan meneliti dampak komposisi lanskap, menggunakan keragaman serbuk sari sebagai pengganti sumber daya yang tersedia, dan kondisi cuaca lokal terhadap kelangsungan hidup koloni lebah madu di musim dingin di berbagai lanskap Eropa. Dengan melakukan eksperimen skala besar selama beberapa tahun di sepanjang gradien iklim di Eropa yang mencakup tiga subtipe iklim utama (samudra, benua, dan Mediterania) tetapi juga gradien iklim dan lanskap skala regional, kami bertujuan untuk memberikan pemahaman yang komprehensif tentang hubungan kausal antara keragaman bunga, cuaca lokal, dan kelangsungan hidup koloni lebah madu di musim dingin. Jika kelangsungan hidup di musim dingin dipengaruhi oleh keragaman serbuk sari yang dikumpulkan oleh lebah pengumpul, maka kami berharap bahwa lanskap di sekitar tempat pemeliharaan lebah dapat memudahkan atau mengintensifkan upaya koloni lebah madu untuk berhibernasi, tergantung pada komposisi habitat. Kondisi cuaca lokal juga dimasukkan sebagai parameter penting dalam kerangka kerja kami karena suhu dan frekuensi presipitasi selama musim mencari makan serta musim dingin dapat memengaruhi kelangsungan hidup koloni lebah madu.
2 BAHAN DAN METODE
2.1 Studi wilayah dan desain eksperimen
Studi kami dilakukan di tiga negara—Prancis, Jerman, dan Yunani—yang termasuk dalam wilayah iklim berbeda dari klasifikasi iklim Köppen-Geiger 1991–2020 (Beck et al., 2023 ), berdasarkan pola curah hujan dan suhu musiman (Gambar 1 ). Ini meliputi: Cfb (iklim maritim, sedang lembap), Dfb (iklim kontinental dan sedang lembap), Csa (iklim Mediterania, hangat), dan Csb (iklim Mediterania sedang). Untuk membangun tempat pemeliharaan lebah eksperimental, kami memilih tiga wilayah iklim untuk setiap negara berdasarkan gradien suhu tahunan rata-rata (MAT) dari tahun 1970 hingga 2000 (Fick & Hijmans, 2017 ), dengan total sembilan wilayah. Di setiap wilayah, kami memilih tiga tempat pemeliharaan lebah dan mengkategorikannya berdasarkan tipe lanskap dominan sebagai semi-alami, pertanian, dan perkotaan (lihat Bagian 2.2 ). Secara total, kami mendirikan 27 tempat pemeliharaan lebah—sembilan di setiap negara—dari musim panas 2021 hingga Juni 2023, yang mencakup dua musim untuk melewati musim dingin dan dua musim mencari makan. Setiap tempat pemeliharaan lebah terdiri dari lima sarang lebah, dikelola dengan praktik pemeliharaan lebah yang terstandarisasi di semua wilayah untuk meminimalkan variabilitas akibat manajemen (lihat Metode Tambahan ). Ratu lebah saudara dari Apis mellifera carnica (Jerman), Apis mellifera macedonica (Yunani), dan Apis mellifera mellifera (Prancis) digunakan di semua sarang lebah, untuk memastikan konsistensi di setiap wilayah.
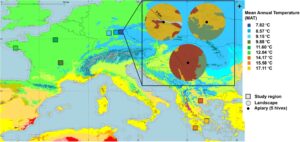
Penelitian ini tidak memerlukan persetujuan dari komite etik hewan karena tidak melibatkan prosedur yang termasuk dalam peraturan percobaan hewan. Selain itu, tidak diperlukan izin atau lisensi khusus untuk kerja lapangan yang dilakukan.
2.2 Komposisi lanskap dan keanekaragaman serbuk sari
Untuk mengukur komposisi lanskap di sekitar setiap tempat pemeliharaan lebah, kami memetakan koordinat geografis setiap tempat pemeliharaan lebah menggunakan QGIS 3.34 (Tim Pengembangan QGIS, 2009 ), menerapkan buffer 2 km untuk mewakili area mencari makan utama bagi lebah madu (Steffan-Dewenter & Kuhn, 2003 ). Data penggunaan lahan diperoleh dari peta CORINE 2018 Land Cover (European Environment Agency dan European Environment Agency, 2019 ), dan tipe habitat yang dihasilkan yang ditemukan di sekitar setiap tempat pemeliharaan lebah dikelompokkan dalam empat kategori utama: semi-alami, pertanian, perkotaan dan lainnya (Tabel S1 ). Data dianalisis untuk menentukan persentase dari setiap kategori utama di setiap lanskap (Gambar S3 ). Keanekaragaman serbuk sari, digunakan sebagai proksi untuk keanekaragaman sumber daya bunga, diukur dengan mengumpulkan sampel serbuk sari setiap bulan dari Maret hingga November 2022, dengan memasang perangkap serbuk sari selama 48 jam di pintu masuk dua sarang untuk setiap tempat pemeliharaan lebah. Secara total, kami menyelesaikan tujuh putaran pengambilan sampel selama musim mencari makan di setiap negara. Karena lokasi tempat pemeliharaan lebah tidak berubah antara tahun 2021 dan 2022, kami berasumsi bahwa keanekaragaman serbuk sari yang dicari di lanskap tertentu akan serupa antara 2 tahun tersebut. Sampel ditimbang, dikeringkan beku selama 24 jam, dan selanjutnya digunakan untuk identifikasi spesies tanaman menggunakan DNA metabarcoding, mengikuti protokol dari Sickel et al. ( 2015 ).
2.3 Data cuaca
Kami mengukur suhu lingkungan lokal pada interval 5 menit di setiap lokasi peternakan lebah, menggunakan sensor suhu digital yang ditempatkan di bawah koloni (sekitar 20–50 cm di atas tanah). Kami mengambil data presipitasi untuk setiap peternakan lebah menggunakan paket iklim R (Czernecki et al., 2020 ) dan memilih stasiun meteorologi yang dekat dengan lokasi yang diinginkan. Kami kemudian menghitung suhu rata-rata dan jumlah kejadian presipitasi (mm) untuk setiap bulan. Untuk analisis selanjutnya, kami menetapkan setiap bulan musim tertentu dan menghitung suhu rata-rata dan jumlah kejadian presipitasi untuk setiap musim. Musim dibagi menjadi cuaca musim semi (Maret, April dan Mei), cuaca musim panas (Juni, Juli dan Agustus), cuaca musim gugur (September, Oktober dan November) dan cuaca musim dingin (Desember, Januari dan Februari). Selain itu, untuk setiap musim, kami juga menjumlahkan jumlah hari yang cocok untuk terbang dan mencari makan lebah madu, dengan suhu rata-rata di atas 10 derajat Celsius dan total presipitasi di bawah 3 mm (Calovi et al., 2021 ; Hatjina et al., 2014 ).
2.4 Analisis statistik
Kami menggunakan DAG untuk mengonseptualisasikan hubungan kausal antara komposisi lanskap, keanekaragaman bunga, kondisi cuaca, dan keberhasilan lebah madu melewati musim dingin. Deskripsi metode untuk DAG disajikan dalam Metode Tambahan (teks dan Gambar S1A ). Analisis statistik kami terdiri dari dua bagian. Pertama, kami tertarik mempelajari bagaimana komposisi lanskap memediasi keanekaragaman serbuk sari yang dicari di setiap tempat pemeliharaan lebah selama musim-musim sebelum musim dingin (Model A). Komposisi lanskap dipelajari dengan menggunakan persentase kategori lanskap utama (seminatural dan pertanian) dalam radius 2 km di sekitar tempat pemeliharaan lebah. Kami mengecualikan persentase area perkotaan dari analisis karena berkorelasi negatif dengan area yang ditempati oleh lahan pertanian (Gambar S2 ). Selain itu, area yang ditempati oleh pertanian cenderung menjadi proksi yang memadai dan dipelajari secara luas untuk mewakili gradien penyederhanaan lanskap (Martin et al., 2019 ). Keanekaragaman serbuk sari (indeks Shannon) diperoleh dari paket vegan (Oksanen et al., 2022 ) dan dihitung dengan mengelompokkan data berdasarkan tempat pemeliharaan lebah untuk setiap tanggal pengumpulan. Berdasarkan bulan pengumpulan, kami mengklasifikasikan sampel kami ke dalam tiga periode utama: serbuk sari musim semi (Maret, April, dan Mei), serbuk sari musim panas (Juni, Juli, dan Agustus), dan serbuk sari musim gugur (September, Oktober, dan November). Pada bagian kedua analisis, kami memodelkan kelangsungan hidup koloni lebah madu di musim dingin dalam hal variabel paparan yang ditentukan oleh DAG (Model B). Karena suhu dan curah hujan secara bersama-sama memengaruhi koloni lebah madu dengan membatasi aktivitas mencari makan dan sangat berkorelasi antar musim (Gambar S2 ), kami memilih untuk menggunakan hari-hari mencari makan sebagai variabel iklim utama dalam penelitian kami. Kami menjumlahkan hari-hari mencari makan di musim semi, musim panas, dan musim gugur untuk mendapatkan jumlah total hari mencari makan selama musim tanam untuk setiap koloni, sementara hari-hari mencari makan di musim dingin dipertimbangkan secara terpisah. Untuk menggambarkan pengaruh sumber daya mencari makan yang tersedia terhadap kelangsungan hidup di musim dingin, kami memanfaatkan keanekaragaman serbuk sari di musim-musim sebelum hibernasi. Keanekaragaman sumber daya bunga sepanjang musim digunakan dalam kasus ini untuk menentukan pengaruh komposisi lanskap sekitarnya terhadap koloni yang berhibernasi.
Untuk kedua model, kami memasang model campuran linear umum (GLMM), menggunakan distribusi Gaussian untuk Model A dan distribusi binomial (‘kelangsungan hidup selama musim dingin’ = 1, ‘kegagalan selama musim dingin’ = 0) dalam Model B. Efek tetap yang disertakan dalam Model A adalah: musim, negara, persentase area semi-alami ( Semi natural ), persentase area pertanian ( Agriculture ). Untuk memperhitungkan struktur data hierarkis, interaksi juga disertakan dalam model ( Agriculture : season , Semi natural : season , country : season ). Prediktor tetap dalam Model B terdiri dari jumlah hari mencari makan selama musim tanam ( fdays_growing ), jumlah hari mencari makan di musim dingin ( fdays_winter ), keanekaragaman musiman sampel serbuk sari, negara dan tahun. Selain itu, kami menyertakan istilah interaksi antara negara dan hari mencari makan di musim tanam dan musim dingin. Perbedaan kelangsungan hidup antara negara dan tahun dimodelkan dalam GLMM terpisah, dengan ‘negara’ dan ‘tahun’ sebagai prediktor tetap.
Semua analisis statistik diimplementasikan menggunakan R v.4.3.1 (Tim Inti R, 2023 ). Kedua model memperhitungkan intersepsi acak untuk setiap kombinasi wilayah dan peternakan lebah dan dipasang menggunakan paket glmmTMB (Brooks et al., 2017 ). Paket DHARMa (Hartig, 2022 ) memungkinkan kami untuk mengevaluasi kinerja model dan mengidentifikasi potensi penyimpangan dari asumsi model, seperti homoskedastisitas dan kenormalan residual; kami tidak menemukan penyimpangan signifikan dari asumsi model. Koefisien model dari GLMM dihasilkan menggunakan paket broom.mixed (Bolker & Robinson, 2022 ) dan emmeans (Lenth, 2023 ). Grafik dihasilkan menggunakan paket R ggplot (Wickham, 2016 ) dan ggeffects (Lüdecke, 2018 ).
3 HASIL
3.1 Hari-hari mencari makan di berbagai wilayah iklim
Kami menemukan hubungan yang jelas antara jumlah hari mencari makan dan MAT di tiga wilayah iklim (Gambar 2 ). Peternakan lebah yang terletak di wilayah iklim kontinental mengalami jumlah hari mencari makan terendah—59 hingga 178 hari selama musim tanam dan antara 1 dan 6 hari selama musim dingin—sementara hari mencari makan di iklim Mediterania berkisar antara 119 dan 238 hari selama musim tanam dan hingga 63 hari mencari makan selama musim dingin.
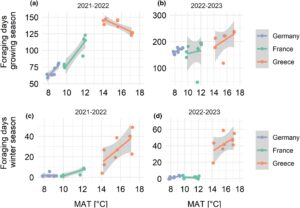
3.2 Keanekaragaman serbuk sari dan komposisi lanskap
Sebanyak 358 sampel serbuk sari dikumpulkan dari 27 lokasi selama musim tanam tahun 2022, di mana total 408 spesies tanaman unik diidentifikasi (Lampiran A ). Kami menemukan bahwa komposisi sumber daya bunga yang digunakan oleh koloni berubah sepanjang musim di berbagai lanskap dan negara, pada tingkat famili (Lampiran A , Gambar A1 ). Kami juga mengamati bahwa persentase tinggi (dari 45% hingga 63%) serbuk sari yang diambil dari tanaman liar di tiga musim (Lampiran A , Gambar A2 ).
Dalam Model A (model komposisi lanskap), keragaman serbuk sari sangat berbeda antar musim (Musim semi, Est. = 0,49, CI = 0,15–0,83, nilai- p < 0,05) tetapi tidak di antara negara-negara pengumpulan (Gambar S4 ). Dengan analisis perbandingan berpasangan, kami menemukan keragaman yang jauh lebih tinggi di musim semi (Est. = 0,37, CI = 0,24–0,49, nilai- p < 0,05) dan musim panas (Est. = 0,33, CI = 0,19–0,45, nilai- p < 0,05) dibandingkan dengan musim gugur. Tidak ada interaksi yang signifikan antara negara dan musim (lihat Tabel S2 untuk estimasi perbandingan berpasangan). Ketika menilai pengaruh komposisi lanskap pada keanekaragaman serbuk sari, model tersebut mengungkap efek positif dari area pertanian (Est. = 0,005, CI = 0,002–0,009, nilai- p < 0,05), sementara interaksi antara musim semi dan area pertanian memiliki efek negatif (Est. = 0,005, CI = 0,002–0,009, nilai- p < 0,05). Namun, tren marginal yang diestimasikan memberikan pandangan yang lebih bernuansa di seluruh musim: di musim panas, area semi-alami berasosiasi positif dengan keanekaragaman serbuk sari yang lebih tinggi dibandingkan dengan area pertanian, sementara di musim gugur, area pertanian menunjukkan efek positif pada keanekaragaman serbuk sari (Gambar 3 ). Selama musim semi, tidak ada efek signifikan dari komposisi lanskap pada keanekaragaman serbuk sari. Keanekaragaman serbuk sari meningkat di tempat pemeliharaan lebah dengan meningkatnya persentase area semi-alami selama musim panas, sementara di musim gugur, meningkatnya keberadaan area pertanian mendukung keanekaragaman serbuk sari yang lebih tinggi (Gambar 4 ).
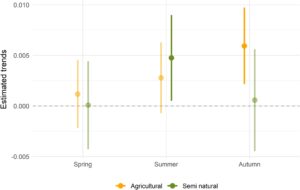
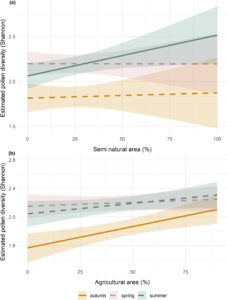
3.3 Bertahan hidup di musim dingin
Rata-rata tingkat kelangsungan hidup musim dingin yang dihitung untuk setiap negara antara 2 tahun bervariasi dari 53,84% hingga 97,77% (Tabel S3 ). Ketika memodelkan kelangsungan hidup koloni lebah madu di berbagai negara, efek marjinal yang dirata-ratakan antar tahun tidak signifikan (Gambar S5 ). Uji post hoc malah menunjukkan tingkat kelangsungan hidup yang lebih tinggi di Jerman dibandingkan dengan Prancis (Est. = 4,02, CI = 1,29–6,75, nilai- p < 0,05) dan Yunani (Est. = −3,00, CI = −5,68 hingga −0,3, nilai- p
< 0,05; Tabel S4 ) mungkin karena tingkat kelangsungan hidup koloni yang tinggi di Jerman pada tahun kedua musim dingin.
Keanekaragaman serbuk sari pada musim semi dan musim gugur muncul sebagai prediktor utama kelangsungan hidup musim dingin pada koloni lebah madu (Gambar S6 ). Secara khusus, keanekaragaman serbuk sari yang tinggi pada musim semi (Est = 2,63, CI = 0,91–4,34) dan pada musim gugur (Est. = 2,34, CI = 0,32–4,36) secara signifikan meningkatkan kelangsungan hidup musim dingin. Probabilitas kelangsungan hidup meningkat dari 19% menjadi 93% dengan peningkatan keanekaragaman serbuk sari musim semi dari 1,8 menjadi 3,4, sementara pada musim gugur peningkatan indeks Shannon dari 0,80 menjadi 3,40 berarti peningkatan 85% dalam tingkat kelangsungan hidup (Gambar 5 ).
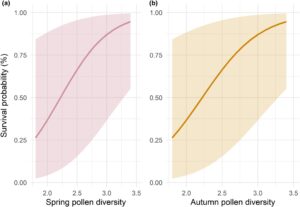
Sebaliknya, hari-hari mencari makan selama musim tanam dan musim dingin tidak memiliki efek signifikan pada kelangsungan hidup di musim dingin (Gambar S6 ). Meskipun demikian, kami mengamati tren positif untuk mencari makan selama musim tanam dan tren negatif untuk hari-hari mencari makan selama musim dingin pada tingkat kelangsungan hidup koloni (Gambar S7 ).
4 DISKUSI
Studi skala besar kami tentang kelangsungan hidup koloni lebah madu di musim dingin menunjukkan bahwa keanekaragaman bunga, khususnya di musim semi dan musim gugur, memainkan peran penting dalam keberhasilan melewati musim dingin. Selain itu, kami mengungkap dampak komposisi lanskap pada preferensi mencari makan dengan efek positif habitat semialami pada keanekaragaman serbuk sari selama musim panas dan habitat pertanian selama musim gugur. Pendekatan ini sangat berguna untuk membandingkan variasi musiman dalam sumber serbuk sari dan pengaruhnya terhadap kelangsungan hidup di musim dingin. Berlawanan dengan harapan kami, kami tidak menemukan pengaruh cuaca yang signifikan terhadap keberhasilan bertahan hidup di musim dingin.
4.1 Variasi musiman keanekaragaman serbuk sari di lanskap dengan komposisi berbeda
Kami mengamati variasi musiman keanekaragaman serbuk sari dan efek komposisi lanskap sekitarnya pada sumber daya bunga. Keanekaragaman secara signifikan lebih tinggi di musim semi (Maret hingga Mei), tetapi selama periode ini, tidak ada perbedaan antara lanskap dengan komposisi habitat yang berbeda. Ini menunjukkan bahwa bahkan di lanskap dengan meningkatnya kehadiran gangguan manusia, terutama pada saat persentase tinggi tanaman berbunga massal diharapkan (lihat juga Harris et al., 2024 ), lebah madu bertujuan untuk mendiversifikasi pencarian makan mereka. Di musim semi, tanaman yang paling umum di lanskap pertanian adalah biji minyak—terutama di Jerman dan Prancis (Wittkop et al., 2009 ). Namun, di iklim Mediterania seperti Yunani, kebun buah juga banyak terdapat di musim semi (Liolios et al., 2015 ; Statistik—ELSTAT, nd ). Juga, selama musim ini, ada tingkat intens pemeliharaan induk, dan seperti yang dipelajari sebelumnya, kami berharap bahwa lebah madu memenuhi kebutuhan mereka untuk protein dengan mencari makan pada keanekaragaman tanaman yang lebih tinggi (Mattila & Otis, 2006 ). Selama musim panas (Juni hingga Agustus), ketika ketersediaan serbuk sari biasanya menurun (Satta et al., 2024 ; Steffan-Dewenter & Kuhn, 2003 ), kami menemukan bahwa peningkatan area habitat semi-alami memiliki efek positif pada keanekaragaman serbuk sari. Temuan ini sejalan dengan Danner et al. ( 2016 ), yang mengamati bahwa jarak mencari makan di musim panas secara keseluruhan berkurang dengan adanya habitat semi-alami di dekat sarang. Menariknya, di akhir musim tanam (September hingga November), lanskap dengan persentase lahan pertanian yang lebih tinggi lebih kuat terkait dengan keanekaragaman serbuk sari, berbeda dengan penelitian Malagnini et al. ( 2022 ), yang hasilnya menunjukkan bahwa habitat semi-alami selama waktu ini lebih penting dalam menentukan keanekaragaman. Di tempat pemeliharaan lebah kami, mayoritas spesies serbuk sari yang dikenali berasal dari tanaman liar (Lampiran A ), yang menunjukkan bahwa mungkin ada kontribusi signifikan dari tepi ladang, jalur bunga, dan pagar tanaman di sekitar ladang yang dibudidayakan (lihat juga Jachuła et al., 2022 ). Namun, hipotesis ini harus ditafsirkan dengan hati-hati karena kami tidak mengukur struktur yang disebutkan di lanskap kami. Namun demikian, selain variabilitas musiman, dalam penelitian ini, kami mengamati bahwa lanskap yang didominasi oleh lahan pertanian dapat mendukung keragaman tanaman yang tinggi yang dibutuhkan untuk sarang selama waktu sebelum hibernasi. Hal tersebut di atas memperkuat gagasan bahwa spesies yang diteliti kurang sensitif terhadap gangguan antropologis dibandingkan dengan penyerbuk lain, yang sebaliknya membutuhkan kedekatan dengan habitat alami untuk tumbuh subur (Biegerl et al., 2025 ; Ricketts et al., 2008) .). Interaksi positif antara keragaman serbuk sari musim gugur dan lanskap dengan persentase habitat pertanian yang lebih tinggi menyoroti kemungkinan perbedaan dalam preferensi mencari makan di antara penyerbuk dan menunjukkan bahwa lanskap pertanian di wilayah studi kami menyediakan sumber serbuk sari yang agak beragam untuk koloni lebah madu di musim gugur. Ini menunjukkan bahwa koloni lebah madu yang dikelola kurang bergantung pada keberadaan area semi-alami daripada penyerbuk liar lainnya (Biegerl et al., 2025 ). Ini menunjukkan, dalam hal manajemen pemeliharaan lebah, bahwa koloni lebah madu di akhir musim panas dan musim gugur dapat memperoleh manfaat dari lanskap pertanian dengan sumber daya bunga yang beragam dan tidak secara khusus membutuhkan kehadiran yang tinggi ketika sebagian besar spesies tanaman liar di area alami dan semi-alami telah menyelesaikan fenologi pembungaannya. Implementasi skema agri-lingkungan yang mencakup ladang bunga dan penanaman sela dengan campuran bunga yang beragam dapat berkontribusi untuk mengisi kesenjangan sumber daya akhir musim dan harus dipromosikan oleh Kebijakan Pertanian Bersama Uni Eropa (CAP) (Boetzl et al., 2021 ).
4.2 Pengaruh keanekaragaman bunga dan cuaca terhadap kelangsungan hidup di musim dingin
Meskipun komposisi lanskap telah dipertimbangkan dalam studi tentang musim dingin lebah madu (Kuchling et al., 2018 ; Requier et al., 2017 ; Rutschmann et al., 2022 ), keanekaragaman bunga tidak pernah dikaitkan secara langsung dengan kelangsungan hidup di musim dingin. Dalam studi kami, kami dapat menetapkan hubungan yang jelas antara keanekaragaman serbuk sari yang dikumpulkan pada musim semi dan musim gugur dan kelangsungan hidup koloni di musim dingin yang lebih tinggi.
Penelitian sebelumnya melaporkan bahwa ketersediaan serbuk sari selama musim semi berkontribusi pada pemeliharaan lebih banyak lebah pekerja dan karenanya mendorong ukuran koloni dan hasil madu yang lebih besar di akhir musim (Hatjina et al., 2014 ). Seperti yang disarankan oleh Requier et al. ( 2017 ), penurunan serbuk sari musim semi yang diburu dapat menyebabkan efek lanjutan, yang mengarah ke koloni dengan jumlah lebah dewasa yang berkurang dan karenanya dengan tingkat kelangsungan hidup musim dingin yang lebih rendah. Penelitian kami menunjukkan bahwa keragaman serbuk sari di musim semi adalah salah satu pendorong utama kelangsungan hidup selama musim dingin. Ini menyoroti bahwa tidak hanya massa serbuk sari yang diburu tetapi juga keragaman makanan memainkan peran penting dalam keberhasilan kelangsungan hidup musim dingin, mungkin dengan meningkatkan kesehatan koloni di awal musim mencari makan. Demikian pula, keragaman serbuk sari musim gugur sangat berkorelasi dengan peningkatan kemungkinan kelangsungan hidup musim dingin. Pemberian serbuk sari selama musim gugur, yang mendahului pembentukan kelompok musim dingin, memelihara lebah musim dingin yang memiliki peran penting dalam termoregulasi dan kelangsungan hidup kelompok pada periode dingin berikutnya (Southwick, 1991 ). Selain itu, pada bulan-bulan menjelang waktu tanpa tanaman berbunga dan mencari makan, serbuk sari disimpan di dalam sarang dan hanya dikonsumsi saat pemeliharaan induk dilanjutkan, sehingga penting untuk pengembangan generasi lebah musim semi berikutnya. Namun, keanekaragaman serbuk sari yang lebih tinggi, dibandingkan dengan serbuk sari dengan nilai gizi yang lebih tinggi (dalam hal persentase protein), tidak pernah secara langsung dikaitkan dengan peningkatan kesehatan lebah individu atau pengembangan koloni yang lebih baik (Pasquale et al., 2013 ). Namun, seperti yang dicatat sebelumnya oleh Alaux et al. ( 2010 ), manfaat dari diet serbuk sari polifloral dan berpotensi dari sumber yang sangat beragam, terletak pada penyediaan lebah dengan komposisi senyawa asam amino yang lebih seimbang dan nutrisi lain yang penting untuk perkembangan dan kesehatan larva lebah madu. Keragaman serbuk sari musim panas sebaliknya tidak relevan dalam hal kelangsungan hidup dalam penelitian kami, bahkan jika penting untuk menyebutkan bahwa kesinambungan sumber daya sepanjang musim dapat menjadi penentu untuk pemeliharaan koloni yang kuat dan sehat.
Sebaliknya, cuaca tampaknya tidak memengaruhi kelangsungan hidup koloni selama musim dingin secara signifikan dalam penelitian kami. Meskipun kami mengamati tren yang menunjukkan bahwa jumlah hari mencari makan dapat memengaruhi kelangsungan hidup selama musim dingin secara negatif, efek ini tidak signifikan secara statistik. Hari-hari musim dingin yang lebih ringan juga dapat bermanfaat bagi kesehatan keseluruhan kelompok musim dingin karena memungkinkan lebah musim dingin untuk melakukan penerbangan higienis dan pengambilan air secara teratur (Becsi et al., 2021 ). Kurangnya korelasi dapat dikaitkan dengan durasi percobaan yang terbatas, dengan data musim dingin selama 2 tahun, karena variasi temporal dalam kondisi lingkungan atau proses biologis mungkin tidak sepenuhnya tertangkap. Sementara kami bertujuan untuk mengatasi keterbatasan ini dengan sejumlah besar replikasi spasial dan cakupan geografis yang luas dalam desain penelitian kami, pendekatan ini tidak dapat sepenuhnya menggantikan replikasi lintas waktu. Penelitian masa depan yang dilakukan dalam periode yang lebih lama akan membantu untuk mengonfirmasi pola yang kami amati dan memberikan pemahaman yang lebih komprehensif tentang sistem dalam berbagai kondisi temporal. Jika tren yang ditemukan dalam studi kami dikonfirmasi, tren tersebut akan menunjukkan kemampuan hebat lebah madu musim dingin untuk beradaptasi dengan berbagai kondisi lingkungan dan efisiensinya untuk mengatasi peristiwa cuaca ekstrem (Stabentheiner et al., 2021 ). Demikian pula, kami tidak mengamati efek signifikan dari hari mencari makan selama musim tanam terhadap kelangsungan hidup musim dingin, meskipun ada tren positif. Penggunaan hitungan hari mencari makan mungkin tidak sepenuhnya menangkap kompleksitas stresor terkait cuaca, dan pelacakan dinamika koloni yang lebih rinci, seperti pencatatan berat dan aktivitas terbang otomatis, dapat memberikan wawasan tambahan tentang bagaimana perilaku mencari makan musiman memengaruhi kesehatan koloni jangka panjang (McMinn-Sauder et al., 2024 ; Meikle et al., 2018 ).
5 KESIMPULAN
Meskipun tingkat Varroa dan ukuran koloni sering dianggap sebagai penyebab utama runtuhnya koloni musim dingin, temuan kami menekankan pentingnya memasukkan sumber daya bunga dan kondisi cuaca dalam kerangka yang lebih luas untuk mempelajari kelangsungan hidup lebah madu selama musim dingin. Meskipun hasil kami mengenai cuaca tidak meyakinkan, hal itu tetap menjadi faktor penting untuk penelitian lebih lanjut, karena pola cuaca lokal tahunan dapat secara signifikan memengaruhi perkembangan koloni dan eksploitasi sumber daya bunga. Keragaman sumber daya yang dikumpulkan oleh para pengumpul makanan selama periode-periode penting—musim semi, saat koloni berkembang, dan musim gugur, saat mereka bersiap untuk musim dingin—sangat penting, karena data kami menunjukkan bahwa hal itu dapat secara signifikan meningkatkan kelangsungan hidup koloni lebah madu. Hal ini menunjukkan bahwa pemantauan sumber daya bunga dan peningkatan keragaman tanaman menarik di sekitar tempat pemeliharaan lebah merupakan strategi praktis untuk meningkatkan kelangsungan hidup lebah madu selama musim dingin. Studi ini juga menyoroti peran penting area pertanian, yang sudah dipengaruhi oleh berbagai tingkat pengelolaan manusia. Oleh karena itu, penerapan skema agri-lingkungan yang mempromosikan spesies tanaman yang menarik bagi lebah madu—seperti membuat ladang dan jalur bunga, mengelola tepi ladang, dan mengoptimalkan praktik pemotongan rumput—dapat diadopsi secara luas (Boetzl et al., 2021 ). Secara keseluruhan, hasil yang disajikan dalam studi ini menunjukkan bahwa menjaga penyediaan sumber daya bunga yang beragam sepanjang musim tanam dapat meningkatkan tingkat kelangsungan hidup koloni lebah madu selama musim dingin dan berpotensi menguntungkan koloni lebah madu yang hidup di alam liar (Kohl et al., 2023 ) dan penyerbuk lain dengan kebutuhan sumber daya yang sama.


Leave a Reply