
Abstrak
Triticale dan lupin adalah kandidat yang menjanjikan untuk intercropping berkelanjutan dalam sistem pertanian campuran tanaman-ternak input rendah di cekungan Mediterania, di mana mereka dapat ditanam sebagai tanaman tadah hujan selama periode musim gugur-musim dingin. Uji coba lapangan 2 tahun dilakukan di Sardinia, Italia, untuk membandingkan intercropping baris triticale–lupin (IC) dengan triticale sole cropping (TSC) dan lupin sole cropping di bawah empat perlakuan pemupukan: hanya nitrogen (‘N,’ 100 kg ha −1 ), hanya fosfor (‘P,’ 39 kg ha −1 ), nitrogen dan fosfor dikombinasikan (‘NP,’ 100 kg ha −1 N + 39 kg ha −1 P), dan kontrol yang tidak dipupuk (‘0’). Pengukuran termasuk intersepsi radiasi, produksi bahan kering (DM), persentase N, dan serapan N per satuan luas (hasil N) pada berbagai pengambilan sampel, bersama dengan hasil gabah dan N pada saat matang. Sementara kedua spesies berbagi durasi siklus tumbuh yang sama, perkembangan tajuk lupin yang lebih lambat dan tinggi yang lebih pendek membatasi intersepsi radiasi dan produksi DM di sela-sela. Namun, komplementaritas dalam DM dan hasil N diamati di keempat tanggal pengambilan sampel di bawah perlakuan 0 dan P, dengan nilai rasio ekuivalen lahan mencapai puncaknya pada 1,71 di bawah perlakuan 0 dan 1,63 di bawah perlakuan P untuk DM, dan sekitar 2 untuk hasil N. DM yang lebih tinggi pada saat matang menghasilkan hasil gabah yang lebih besar untuk IC dibandingkan dengan TSC di bawah perlakuan 0 (3,9 vs. 3,2 t ha −1 ) dan P (4,3 vs. 3,6 t ha −1 ). IC juga mengungguli TSC dalam hal persentase N DM, terutama pada antesis triticale, dengan perbedaan nyata pada perlakuan 0 (N% = 1,09 untuk IC, 0,79 untuk TSC) dan P (N% = 1,17 untuk IC, 0,83 untuk TSC). Persentase N DM IC yang lebih tinggi ini kemungkinan terkait dengan efisiensi penggunaan radiasi IC yang lebih tinggi dibandingkan dengan TSC di bawah perlakuan pemupukan 0. Lebih jauh, penanaman silang triticale dengan lupin menunjukkan potensi untuk menggantikan pemupukan N tanpa mengorbankan DM atau kandungan N dalam DM dan dalam jerami. Komplementaritas yang diamati tanpa adanya pupuk menunjukkan bahwa penanaman silang ini dapat sangat cocok untuk sistem input rendah. Pendekatan eksperimental membantu menghubungkan manfaat terkait N dari penanaman silang serealia-kacang-kacangan dengan perolehan radiasi dan efisiensi penggunaan radiasi, sambil menyoroti peran penting laju perkembangan tajuk, dan karenanya pilihan kultivar, ketika perbedaan ketinggian cukup besar.
1. PENDAHULUAN
Dengan penggunaan global pupuk nitrogen (N) sebesar 109 Tg tahun -1 (FAOSTAT, 2019 ), estimasi emisi gas rumah kaca dari produksi pupuk N adalah antara 229 dan 545 Tg CO 2 ekuivalen (Jensen et al., 2020 ). Sebaliknya, tidak ada biaya karbon fosil yang terkait dengan layanan ekosistem yang berharga dari fiksasi N simbiosis yang didorong oleh fotosintesis oleh legum (Peoples et al., 2009 ). Di lingkungan Mediterania, sistem pertanian tadah hujan bergantung pada sereal musim dingin sebagai tanaman komersial, sementara legum dapat ditanam baik sebagai tanaman tunggal (SC) dalam rotasi dengan sereal atau ditanam secara sela dengan sereal. Bahasa Indonesia: Setelah terjadi penurunan tajam dalam budidaya kacang-kacangan sebagai tanaman pakan tunggal selama tahun 1960-an—yang disebabkan oleh pemisahan spasial produksi ternak dan pakan di wilayah pertanian (Monti et al., 2019 )—penanaman tumpang sari serealia–kacang-kacangan telah menarik perhatian untuk sistem pertanian input rendah dan wilayah marjinal (Jensen et al., 2020 ; Księżak et al., 2023 ), seperti sistem pertanian tanaman campuran–ternak petani kecil di cekungan Mediterania.
Interaksi kompetitif dan perolehan N komplementer dalam intercropping memungkinkan serealia memperoleh sumber daya N tanah yang sangat tinggi (Fletcher et al., 2016 ; Jensen et al., 1996 ). Peningkatan penyerapan N oleh serealia ini biasanya mendorong legum biji-bijian untuk mengimbanginya dengan mengikat lebih banyak N atmosfer 2 (Hauggaard-Nielsen et al., 2008 , 2009 ; Rodriguez et al., 2020 ), yang merupakan keuntungan utama dari intercropping serealia-legum dibandingkan rotasi legum-sereal (Fletcher et al., 2016 ). Keuntungan ini terbukti terutama dalam kondisi ketersediaan N tanah yang rendah, di mana intercropping tidak hanya meningkatkan hasil tetapi juga meningkatkan kandungan protein biji-bijian serealia. Peningkatan efisiensi penggunaan N dalam intercropping biji-kacangan-sereal diperkirakan berpotensi mengurangi kebutuhan global untuk pupuk N sintetis sekitar 26% (Jensen et al., 2020 ).
Manfaat lain dari tanaman sela serealia-kacang-kacangan meliputi berkurangnya serangan gulma (Corre-Hellou et al., 2011 ), peningkatan kualitas pakan ternak (Księżak et al., 2023 ), peningkatan nilai pra-tanam (Monti et al., 2019 ; Pappa et al., 2012 ), dan stabilitas hasil panen yang lebih baik (Hauggaard-Nielsen et al., 2009 ; Jensen, 1996 ). Keuntungan ini berpotensi mengatasi tantangan daya saing gulma yang rendah dan hasil panen yang rendah dan tidak stabil yang sering kali memengaruhi SC kacang-kacangan (Bedoussac et al., 2015 ).
Lebih jauh lagi, pemulihan campuran kacang-kacangan serealia-biji-bijian dalam sistem pertanian Mediterania yang tadah hujan dan input rendah—di mana serealia mendominasi—mendorong diversifikasi genetik dan intensifikasi ekologi dengan meningkatkan kontribusi siklus alami dan aliran sumber daya dalam sistem pertanian. Di daerah rawan kekeringan di Italia Selatan, penyertaan kacang-kacangan biji-bijian dalam rotasi dengan gandum durum, tanaman komersial tradisional, merupakan praktik yang sudah lama ada (Monti et al., 2019 ). Ketika ditanam secara tumpang sari, serealia dan kacang-kacangan dapat ditanam dan dipanen secara bersamaan menggunakan peralatan pertanian tradisional.
Selama dekade terakhir, ada minat yang meningkat pada penanaman campur serealia-kacang-kacangan untuk pertanian berkelanjutan, tetapi sebagian besar penelitian berfokus pada jagung dan gandum di antara serealia, dan kedelai dan kacang polong di antara kacang-kacangan (Landschoot et al., 2024 ). Sementara penanaman campur tanaman utama ini menawarkan banyak keuntungan dibandingkan penanaman tunggal (Blessing et al., 2022 ), hal itu tidak berkontribusi secara signifikan untuk meningkatkan diversifikasi tanaman. Sebaliknya, triticale dan lupin kurang mendapat perhatian, dengan hanya 153 dan 253 makalah, masing-masing, dari 4732 makalah tentang penanaman campur serealia-kacang-kacangan, sebagaimana diidentifikasi melalui analisis bibliometrik yang dikombinasikan dengan penambangan teks dan pemodelan topik (Landschoot et al., 2024).
Triticale (× Triticosecale Wittmack) utamanya digunakan untuk pakan ternak, menawarkan nilai energi yang sebanding dengan serealia lain yang ditanam di musim dingin, baik digunakan segar maupun sebagai silase saat matang atau saat matang seperti lilin susu (Faccini et al., 2023 ). Kebutuhan inputnya yang lebih rendah, khususnya tidak adanya kebutuhan irigasi, membuat diet berbasis triticale sekitar 15% lebih hemat air daripada diet berbasis jagung (Faccini et al., 2023 ). Selain itu, triticale telah disarankan sebagai serealia yang cocok untuk ditanam sela dengan kacang-kacangan karena daya saingnya yang lebih rendah dibandingkan dengan serealia lainnya (Ross et al., 2005 ).
Di antara kacang-kacangan, lupin adalah tanaman yang menjanjikan, menawarkan produk yang sangat mudah dicerna, hasil tinggi, dan kaya protein—sifat yang diinginkan untuk memberi makan hewan monogastrik dan ruminansia (van Barneveld, 1999 ). Benih spesies lupin asli Eropa ( Lupinus albus L., Lupinus luteus L. dan Lupinus angustifolius L.) adalah alternatif yang valid untuk kedelai sebagai sumber protein dalam nutrisi hewan, dengan kandungan protein berkisar antara 35% hingga 40%, sebanding dengan kedelai dan lebih tinggi dari kacang polong dan kacang faba (Norton et al., 1985 ). Lupin putih ( L. albus L.) dapat berhasil disilangkan sebagai tanaman utuh (Fraser et al., 2005 ). Seperti triticale, lupin ditanam sebagai tanaman tadah hujan selama periode musim gugur-musim dingin di lingkungan Mediterania, membuatnya lebih ekonomis daripada kedelai, yang memerlukan irigasi karena hanya dapat ditanam di musim semi dan musim panas. Selain itu, lupin mengikat N 2 atmosfer dalam jumlah yang signifikan , dengan tingkat fiksasi rata-rata 75% (Carton et al., 2020 ), dan ketika ditanam secara tumpang sari dengan gandum, lupin melepaskan fosfor yang cukup untuk mendukung pertumbuhan tanaman pendampingnya (Marschner et al., 1986 ).
Ciri-ciri ini menjadikan triticale dan lupin sebagai kandidat yang menjanjikan untuk penanaman tumpang sari yang berkelanjutan. Akan tetapi, hanya ada sedikit penelitian yang menganalisis penanaman tumpang sari triticale–lupin dalam uji lapangan (Azo et al., 2012 ; Carton et al., 2020 ; Górski & Płaza, 2024 ; Księżak & Staniak, 2013 ), dan belum ada yang meneliti jenis penanaman tumpang sari ini di iklim Mediterania.
Pemberian nutrisi, baik organik maupun anorganik, dapat memengaruhi dinamika persaingan dalam tanaman sela serealia-kacang-kacangan. Baik perolehan N maupun pembagian N antara komponen serealia dan kacang-kacangan diubah oleh waktu dan tingkat pemupukan N (Naudin et al., 2010 ), dan oleh kelembapan tanah dan karenanya oleh jumlah dan distribusi curah hujan. Selain itu, penanaman sela dapat meningkatkan ketersediaan hayati nutrisi seperti P, Fe, Zn, dan Mn, meningkatkan penyerapannya oleh spesies yang ditanam sela (Li et al., 2014 ). Fosfor, khususnya, merupakan elemen penting untuk fiksasi N simbiosis pada kacang-kacangan (Zhong et al., 2023 ).
Percobaan ini bertujuan untuk menganalisis tanaman sela triticale–lupin yang ditanam dengan empat tingkat pemupukan selama uji coba lapangan selama 2 tahun di lingkungan Mediterania, dengan tujuan sebagai berikut:
- Menyelidiki komplementaritas antara kedua spesies dan bagaimana hierarki kompetitif berkembang seiring waktu terkait penangkapan dan penggunaan radiasi;
- Tentukan apakah penanaman sela triticale–lupin merupakan alternatif yang layak untuk triticale di SC
2 BAHAN DAN METODE
Percobaan dilakukan selama musim tanam 2009–2010 (‘2010’) dan 2011–2012 (‘2012’) di lahan percobaan Santa Lucia, Universitas Sassari (lintang 39°54′N; 8°22′E; ketinggian 15 m dpl). Percobaan tidak dapat dilakukan pada musim 2010–2011 karena curah hujan musim gugur yang tinggi, sehingga tidak memungkinkan untuk mengakses lahan dengan mesin. Tanahnya berupa lempung-lempung yang berasal dari aluvial, dengan kedalaman sekitar 2 m, dengan reaksi sub-alkali yang disebabkan oleh natrium dalam kompleks pertukaran. Tanah ini memiliki kandungan N total (0,06%) dan bahan organik (0,6%) yang rendah, tetapi kaya akan fosfor di lapisan atas 0,4 m. Iklimnya khas Mediterania, dengan curah hujan tahunan rata-rata jangka panjang sebesar 575 ± 139 mm, yang terkonsentrasi terutama antara Oktober dan April, dan dengan curah hujan musim semi yang sangat tidak menentu, rata-rata 137 mm dari Maret hingga Mei. Januari adalah bulan terdingin (suhu rata-rata 9,8 ± 2,5°C) dan Agustus adalah bulan terpanas (suhu rata-rata 24,6 ± 2,8°C).
Perlakuan yang diterapkan terdiri dari tiga sistem tanam dengan empat perlakuan pemupukan. Sistem tanam adalah lupin sebagai tanaman tunggal (LSC), triticale sebagai tanaman tunggal (TSC), dan penanaman sela baris lupin dan triticale (IC). Perlakuan pemupukan adalah: N saja (‘N,’ 100 kg ha −1 N), fosfor saja (‘P,’ 39 kg ha −1 P), nitrogen dan fosfor (‘NP,’ 100 kg ha −1 N + 39 kg ha −1 P), dan kontrol tanpa pupuk (‘0’). Sistem tanam dan perlakuan pemupukan disusun dalam rancangan petak-strip dengan empat ulangan, di mana sistem tanam ditetapkan pada kolom dan perlakuan pemupukan pada baris.
Pra-tanam adalah triticale. Persemaian disiapkan melalui pembajakan dan penggaruan. Pemupukan diberikan dengan tangan pada hari yang sama dengan penggaruan, sesuai dengan spesifikasi perlakuan. Penaburan dilakukan dengan seeder baris pada 25/11 di tahun pertama dan pada 30/11 di tahun kedua, di plot seluas 48,6 m 2 (panjang 18 m, lebar 2,7 m) menggunakan rancangan penggantian (Willey, 1979 ) (Gambar S1 ). Untuk lupin ( L. albus L.), kultivar semi-determinat ‘Ares’ ditabur dengan takaran 25 benih yang dapat berkecambah m −2 , setara dengan dosis benih 100 kg ha −1 , dengan jarak baris 0,45 m di plot SC. Untuk triticale, kultivar musim semi ‘Oceania’ digunakan, ditanam dengan takaran 350 biji berkecambah m −2 , setara dengan takaran benih 180 kg ha −1 , dengan jarak tanam 0,15 m. Di petak tumpang sari, kedua spesies ditanam dalam baris bergantian, dengan jarak tanam 0,225 m, dengan setengah takaran tanam yang digunakan di petak SC, yaitu 90 kg ha −1 untuk triticale dan 50 kg ha −1 untuk lupin.
Setelah penanaman, tanah digiling, dan penyiangan pra-tumbuh dilakukan pada tanggal 26 November pada tahun pertama dan 1 Desember pada tahun kedua, menggunakan 3 L ha −1 produk komersial yang mengandung 31,7% pendimethalin.
2.1 Pengukuran
Dalam kedua tahun tersebut, jumlah tanaman per meter persegi ditentukan segera setelah kemunculan dan lagi mendekati pematangan akhir, ketika jumlah batang dihitung untuk triticale, karena tidak mungkin untuk menghitung tanaman satu per satu. Fenologi dipantau melalui inspeksi plot mingguan, dengan setiap tahap dicatat ketika 75% tanaman dalam plot menunjukkan tahap tersebut. Tahapan fenologi yang dicatat untuk lupin adalah: kemunculan, antesis (bunga pertama terbuka), polong pertama (munculnya polong pertama, panjang 8–10 mm), polong kuning (perubahan warna polong dari hijau menjadi coklat pucat), dan kematangan (kematangan panen, dengan biji pada kelembaban sekitar 12%) (Dracup & Kirby, 1993 ). Untuk triticale, tahapan yang dicatat adalah: kemunculan, booting (DC 49, Zadoks et al., 1974 ), antesis (DC 61), kematangan adonan susu (DC 79–80), dan kematangan panen (DC 92). Pada tahun 2012, tinggi tanaman diukur pada empat kesempatan sebagai jarak dari tanah ke puncak batang utama (lupin) atau ke ligula daun terakhir yang berkembang (triticale). Pada kedua musim, tinggi tanaman total juga diukur pada saat dewasa dengan cara yang sama untuk lupin, sebagai jarak dari ujung tongkol (tidak termasuk awn) ke tanah untuk triticale.
Empat pengambilan sampel biomassa dilakukan selama siklus pertumbuhan 2010, dan enam selama 2012, pada tahap fenologi berikut: pra-antesis lupin (28 Februari 2012), antesis lupin (15 Maret 2010 dan 14 Maret 2012), kemunculan polong lupin pertama (26 Maret 2012), antesis triticale (07 April 2010 dan 11 April 2012), kematangan adonan susu triticale (10 Mei 2010 dan 08 Mei 2012), dan kematangan panen (14 Juli 2010 dan 04 Juli 2012). Ukuran sampel untuk triticale di plot TSC adalah 0,45 m 2 untuk pengambilan sampel pertama, 0,9 m 2 pada saat kematangan, dan 0,3 m 2 untuk pengambilan sampel antara. Untuk plot LSC, semua sampel berukuran 0,9 m 2 , kecuali sampel panen yang berukuran 2,7 m 2 . Ukuran sampel yang sama digunakan di plot IC, tempat lupin dan triticale dikumpulkan secara terpisah. Saat matang, sampel dibagi secara manual menjadi jerami dan bulir, dan bulir dirontokkan dengan tangan untuk mendapatkan gabah. Semua sampel dikeringkan dalam oven pada suhu 80°C selama 48 jam untuk mendapatkan bahan kering (DM) dan selanjutnya digiling dengan penggilingan pakan ternak. Di plot IC, kedua spesies digiling secara terpisah. Semua sampel awalnya dianalisis di laboratorium Regional Breeders’ Association (ARA) di Nuraxinieddu (Oristano) menggunakan teknik Near InfraRed untuk menentukan DM dan kandungan protein kasar. Data kemudian dikenakan uji statistik untuk mengidentifikasi outlier, dan sampel outlier dianalisis ulang menggunakan metode Kjeldahl. Kandungan N dihitung dengan membagi protein kasar dengan 6,25, kemudian dikalikan dengan DM untuk memperkirakan jumlah penyerapan N per satuan luas (hasil N).
Data cuaca (suhu maksimum dan minimum, curah hujan, radiasi matahari, dan kelembaban relatif) dicatat oleh stasiun meteorologi yang berjarak sekitar 300 m dari lapangan.
Radiasi aktif fotosintesis (PAR) yang dicegat oleh tajuk diukur selama jam-jam tengah hari menggunakan SunScan Canopy Analysis System SS1-UM-1.05 (Delta-T Devices Ltd., Cambridge, Inggris). PAR rata-rata di permukaan tanah ( I ) dinilai dengan memposisikan probe tegak lurus dengan arah barisan dan sejajar dengan permukaan tanah di tiga titik sepanjang plot. Pada saat yang sama, Sensor Fraksi Berkas memantau cahaya yang datang di permukaan tajuk ( I 0 ). Fraksi PAR yang dicegat (FIPAR) dihitung sebagai (1 − I / I 0 ). Indeks luas daun (LAI) diperkirakan secara tidak langsung menggunakan instrumen yang sama. Di plot tumpang sari, intersepsi radiasi diukur dengan cara yang sama dengan plot tanaman tunggal, tetapi radiasi yang dicegat oleh setiap spesies dibedakan dengan mengambil serangkaian pembacaan kedua setelah mengeluarkan tanaman lupin dari area pengambilan sampel. Hasil pembacaan ini digunakan untuk menghitung FIPAR triticale di sela-sela, dengan intersepsi lupin diperkirakan sebagai selisih dari total. Enam pengukuran dilakukan pada tahun 2010 dan 15 pada tahun 2012, dari Januari hingga Mei.
2.2 Perhitungan dan statistik
Perkembangan tajuk harian dan pola intersepsi radiasi dihitung dengan cara mencocokkan FIPAR dengan model yang diusulkan oleh Steduto et al. ( 2009 ). Sebuah kurva dibuat untuk setiap kombinasi ‘musim × perlakuan pemupukan × sistem tanam’, kecuali untuk lupin yang ditanam di sela-sela, di mana interpolasi tidak memungkinkan karena nilai yang diamati sangat rendah dan bervariasi. PAR kumulatif yang dicegat (IPAR, MJ m −2 ) pada setiap tanggal pengambilan sampel biomassa dihitung sebagai jumlah IPAR harian, yang diperoleh dengan mengalikan nilai FIPAR harian dengan PAR harian, yang diperkirakan sebesar 50% dari radiasi matahari yang tercatat di stasiun meteorologi. Nilai IPAR kumulatif untuk setiap pengambilan sampel biomassa digunakan untuk memperkirakan efisiensi penggunaan radiasi (RUE, g MJ −1 ), yang dihitung sebagai kemiringan regresi DM versus IPAR. Perbedaan nilai RUE di seluruh perlakuan dalam setiap tahun dibandingkan dengan menganalisis kemiringan regresi menurut Gomez dan Gomez ( 1984 ).
Untuk membandingkan efisiensi SC dan intercropping-nya, digunakan land equivalent ratio (LER; Willey & Osiru, 1972 ). LER menunjukkan luas lahan relatif yang dibutuhkan ketika menanam SC untuk menghasilkan jumlah DM atau gabah atau hasil N yang dihasilkan oleh intercropping dengan proporsi spesies yang sama (Willey & Osiru, 1972 ). LER >1 mengindikasikan keuntungan per area untuk intercropping dibandingkan dengan penanaman tunggal dan dengan demikian meningkatkan penggunaan sumber daya lingkungan. LER dihitung mengikuti Willey dan Osiru ( 1972 ) sebagai jumlah dari LER parsial (PLER) dari dua spesies:
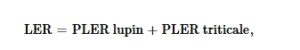
di mana PLER lupin = Y lupin pada tumpang sari/Y lupin pada SC; PLER triticale = Y triticale pada tumpang sari/Y triticale pada SC.
Y tetap untuk sifat-sifat yang LER-nya dihitung, yaitu total DM dan hasil N pada berbagai pengambilan sampel DM, DM gabah dan jerami, dan hasil N pada saat matang.
Analisis statistik dilakukan menggunakan perangkat lunak R (Tim Inti R, 2017 ), dengan paket ‘agricolae’ dan prosedur strip-plot. Kesalahan A, 6 derajat kebebasan (df) (Blok × Sistem penanaman MS) digunakan untuk menguji signifikansi ‘Sistem penanaman,’ Kesalahan B, 9 df (Blok × Pemupukan MS) digunakan untuk menguji signifikansi perlakuan ‘Pemupukan’, dan Kesalahan C, 18 df (Blok × Sistem penanaman × Pemupukan MS) digunakan untuk menguji signifikansi interaksinya. Rata-rata dibandingkan menggunakan uji perbedaan signifikan terkecil (LSD), dan kesalahan standar dan nilai- t yang sesuai diterapkan untuk setiap perbandingan (Gomez & Gomez, 1984 ).
3 HASIL
3.1 Komposisi tanaman sela dan cuaca
Dibandingkan dengan rasio 50:50 yang dimaksudkan untuk tanaman sela, jumlah tanaman triticale yang dihitung di petak IC saat muncul adalah 59% dari jumlah tanaman triticale di petak TSC pada tahun 2010 dan 44% pada tahun 2012 (Tabel S1 ). Sebaliknya, jumlah tanaman lupin di tanaman sela adalah 50% dari jumlah SC pada tahun 2010 dan 61% pada tahun 2012.
Untuk menilai perubahan komposisi IC antara kemunculan dan kematangan, batang triticale (baik yang steril maupun fertil) dan tanaman lupin dihitung saat kematangan. Penurunan kepadatan batang triticale yang signifikan ( p ≤ .05) diamati pada plot TSC dari kemunculan hingga kematangan, lebih besar pada tahun 2010 (dari 356 tanaman m −2 menjadi 293 batang m −2 ) dibandingkan pada tahun 2012 (dari 339 tanaman m −2 menjadi 305 batang m −2 ). Namun, di plot IC, peningkatan yang signifikan pada kepadatan batang triticale diamati pada tahun 2012, meningkat dari 150 tanaman m -2 saat kemunculan menjadi 230 batang m -2 saat dewasa, kemungkinan karena anakan yang intens, sedangkan pada tahun 2010 jumlah batang triticale di IC tetap relatif konstan antara kemunculan dan dewasa (210 tanaman m -2 saat kemunculan dan 202 batang m -2 saat dewasa). Untuk lupin, penurunan yang signifikan ( p ≤ .05) dalam kepadatan tanaman dari kemunculan hingga dewasa diamati di LSC (dari 32 tanaman m −2 menjadi 22 m −2 pada tahun 2010, dan dari 29 tanaman m −2 menjadi 23 m −2 pada tahun 2012) dan plot IC (dari 16 tanaman m −2 menjadi 10 m −2 pada tahun 2010, dan dari 18 tanaman m −2 menjadi 15 m −2 pada tahun 2012).
Dinamika yang berbeda yang diamati dalam dua musim dalam jumlah batang/anakan dari dua spesies antara kemunculan dan kematangan sebagian dapat dikaitkan dengan perbedaan jumlah dan distribusi curah hujan dalam dua musim (Gambar S2 ). Curah hujan musiman (Oktober–Juni) berjumlah 636 mm pada tahun 2010 dan 513 mm pada tahun 2012. Curah hujan musim dingin, terutama pada bulan Januari, secara signifikan lebih tinggi pada tahun 2010 (117 mm dibandingkan dengan 14 mm pada tahun 2012). Mengingat rendahnya evapotranspirasi acuan yang dihitung untuk bulan Januari 2010 (hanya 31 mm), surplus curah hujan (di mana curah hujan melebihi evapotranspirasi) adalah 87 mm. Dikombinasikan dengan tekstur tanah liat, curah hujan yang berlebihan ini mengakibatkan genangan air. Curah hujan pada bulan-bulan kritis April dan Mei, saat perkembangan dan pertumbuhan gabah terjadi, masing-masing mencapai 169 dan 172 mm pada tahun 2010 dan 2012—jauh lebih tinggi daripada rata-rata jangka panjang sebesar 89 mm (berdasarkan data selama 65 tahun). Musim 2012 mengalami suhu minimum dan maksimum yang lebih tinggi dari Oktober hingga Desember dan suhu minimum yang lebih rendah dari Februari hingga Mei. Februari merupakan bulan terdingin pada tahun 2012, dengan suhu bulanan rata-rata 2,1°C, dan suhu serendah 0,3°C selama dekade kedua bulan tersebut. Sebaliknya, Januari merupakan bulan terdingin pada tahun 2010, dengan suhu rata-rata 6,6°C.
3.2 Perkembangan dan tinggi tanaman
Lupin muncul 7 hari lebih lambat dari triticale pada tahun 2010 dan 4 hari kemudian pada tahun 2012 (Gambar S3 ). Antesis lupin (bunga pertama kali mekar) terjadi pada tanggal 15–16 Maret di kedua tahun tersebut, dengan polong pertama terbentuk pada tanggal 23–24 Maret, hampir bersamaan dengan tahap pembentukan sepatu bot triticale. Triticale berbunga rata-rata pada tanggal 7 April tahun 2010 dan 12 April tahun 2012, mencapai tahap kematangan adonan susu sekitar 1 bulan kemudian. Baik antesis triticale maupun kematangan adonan susu terjadi selama fase pertumbuhan polong dan biji lupin, yang berakhir dengan tahap polong kuning, sangat dekat dengan kematangan panen triticale. Kedua spesies tersebut mencapai kematangan panen dalam waktu 3–6 hari satu sama lain, sekitar 6 bulan setelah penaburan.
Tidak ada perbedaan signifikan dalam tinggi tanaman triticale atau tingkat perkembangan yang diamati antara perlakuan pemupukan, atau antara intercrop (IC) dan SC. Triticale mencapai tinggi tanaman akhir yang sama sekitar 108 cm di SC dan IC di kedua tahun. Itu lebih tinggi dari lupin, yang mencapai 90 cm pada tahun 2010 dan 87 cm pada tahun 2012 di SC. Namun, intercropping pada tahun 2012 mengurangi tinggi akhir lupin menjadi 60 cm, karena penghentian perpanjangan mulai sekitar 20-25 Maret, ketika triticale mencapai tahap booting dan mulai memanjang tangkai dengan cepat, bertepatan dengan munculnya polong lupin pertama (Gambar S4 ).
3.3 Intersepsi radiasi, RUE dan akumulasi bahan kering
Pada kedua tahun dan semua perlakuan, intersepsi radiasi maksimum oleh lupin yang ditanam tunggal sebanding dengan triticale tetapi terjadi lebih dari 2 bulan kemudian karena peningkatan linier yang tertunda dalam intersepsi untuk spesies polong-polongan (Gambar 1 ).
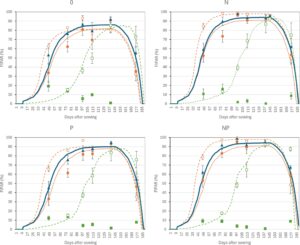
Sebaliknya, lupin di IC menunjukkan intersepsi yang sangat rendah, mungkin karena perawakannya yang lebih pendek, dengan nilai maksimum 15%–20% tercatat sekitar waktu triticale mencapai intersepsi maksimumnya, sekitar 2 bulan setelah disemai.
Semua kombinasi sistem tanam dan perlakuan pemupukan mencapai nilai FIPAR maksimum mendekati atau melebihi 90%, kecuali untuk perlakuan 0 pada tahun 2010, yang berkisar antara 80% dan 90%. Triticale di SC menunjukkan nilai FIPAR yang lebih tinggi daripada intercrop hingga nilai maksimum tercapai, karena kontribusi lupin yang dapat diabaikan terhadap intersepsi intercrop selama periode ini. Perbedaan FIPAR ini menghasilkan nilai IPAR kumulatif yang serupa selama 2 tahun untuk triticale di SC (890 MJ m −2 ), triticale di IC (811 MJ m −2 ) dan intercrop (859 MJ m −2 ). Nilai yang lebih rendah diamati untuk lupin di SC (625 MJ m −2 ) (Gambar S5 ).
Dengan menggunakan data IPAR, RUE dihitung sebagai kemiringan regresi antara DM kumulatif dan IPAR kumulatif (Tabel 1 ).
| Tahun 2010 | Tahun 2012 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| RUE (g MJ -1 ) | Bahasa Inggris | Mungkin > | t | | Interval kepercayaan | RUE (g MJ -1 ) | Bahasa Inggris | Mungkin > | t | | Interval kepercayaan | |||
| L1 | Bahasa Indonesia: L2 | L1 | Bahasa Indonesia: L2 | |||||||
| IC | 1.61 | 0,09 | <.0001 | 1.42 | 1.80 | 2.19 | 0.10 | <.0001 | 1.97 | 2.40 |
| LSC | 1.28 | 0,06 | <.0001 | 1.16 | 1.41 | 1.52 | 0,09 | <.0001 | 1.34 | 1.70 |
| TSC | 1.45 | 0.12 | <.0001 | 1.19 | 1.71 | 2.41 | 0.12 | <.0001 | 2.16 | 2.65 |
| TIKTAK | 1.42 | 0,09 | <.0001 | 1.23 | 1.61 | 2.21 | 0.12 | <.0001 | 1,95 | 2.47 |
Catatan : Nilai RUE dengan interval kepercayaan yang tumpang tindih tidak dapat dianggap berbeda secara signifikan untuk p ≤ .05.
RUE pada umumnya lebih rendah pada tahun 2010 dibanding tahun 2012, terutama untuk TSC (60% dari RUE tahun 2012) dan IC (64% dari RUE tahun 2012), suatu perbedaan yang menjelaskan mengapa DM final tahun 2012 lebih besar dibanding tahun sebelumnya, terutama untuk TSC (+97%) (Gambar 2 dan Tabel S2 ).
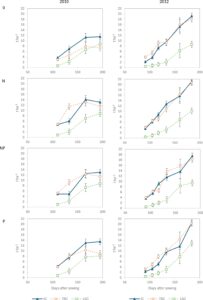
Lupin yang ditanam tunggal memiliki RUE terendah di kedua tahun tersebut, sementara hanya pada tahun 2010 intercrop menunjukkan RUE rata-rata tertinggi, meskipun tidak berbeda secara signifikan dari triticale. Keunggulan RUE intercrop atas RUE triticale yang ditanam tunggal paling menonjol dan signifikan ( p ≤ .05) dalam perlakuan 0 (Tabel S3 ), di mana RUE intercrop adalah 1,58 ± 0,057 g MJ −1 , dibandingkan dengan 1,10 ± 0,037 g MJ −1 untuk triticale di SC. Pada tahun 2012, DM untuk IC dan TSC tidak berbeda secara signifikan, terlepas dari tanggal pengambilan sampel atau perlakuan pemupukan, sementara DM lupin secara konsisten adalah yang terendah (Tabel S2 ). Sebaliknya, pada tahun 2010, IC menghasilkan DM yang lebih tinggi daripada TSC dalam perlakuan 0 dan dalam dua pengambilan sampel terakhir dari perlakuan P. Khususnya, pada kedua tahun tersebut, total DM yang diproduksi oleh IC pada perlakuan 0 tidak berbeda dengan DM yang diproduksi oleh TSC di bawah pemupukan N.
Menurut nilai PLER, lupin terpengaruh secara negatif oleh penanaman tumpang sari dalam hal produksi DM, khususnya pada tahun 2012, ketika PLER-nya secara progresif menurun dari maksimum 0,17 menjadi 0,03, yang menunjukkan bahwa DM lupin hampir menghilang dari plot penanaman tumpang sari (Gambar 3 ). Hal ini mencegah adanya komplementaritas yang signifikan antara spesies, sebagaimana tercermin dalam nilai LER rata-rata yang rendah sebesar 1,02 untuk tahun itu. DM lupin yang rendah disebabkan oleh pengurangan jumlah tanaman per meter persegi dan berat tanaman yang berkurang yang mungkin terkait dengan tinggi tanaman akhir yang secara signifikan lebih rendah daripada di SC. Meskipun demikian, triticale menghasilkan jumlah DM yang sama di SC dan penanaman tumpang sari (dengan PLER rata-rata 0,92), kemungkinan karena anakannya yang produktif, yang mengimbangi berkurangnya area yang ditanami triticale di IC dibandingkan dengan SC. Sebaliknya, pada tahun 2010, PLER triticale sebagian besar melampaui 0,5 pada perlakuan 0 (1,14, 1,31 dan 1,12 dan 1,11 pada empat sampel DM), yang menunjukkan bahwa triticale diuntungkan oleh keberadaan lupin. Komplementaritas ini menghasilkan nilai LER pada perlakuan 0 sebesar 1,59, 1,71 dan 1,56 dan 1,49 pada empat sampel. Nilai LER yang tinggi pada perlakuan P, yang mencapai puncaknya 1,63 pada sampel pertama, terutama didorong oleh PLER lupin, yang menurun dari 0,68 pada sampel pertama menjadi 0,33 pada sampel terakhir.

3.4 Akumulasi Nitrogen
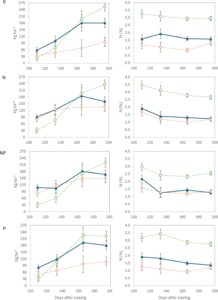
Pada tahun 2012, perbedaan antara IC dan TSC terbatas pada persentase N yang lebih besar di IC dibandingkan dengan TSC, tetapi hanya pada pengambilan sampel pertama. Hal ini merupakan konsekuensi dari keberadaan lupin yang langka di IC selama tahun ini (Tabel S4 ). Sebaliknya, pada tahun 2010, keberadaan lupin di IC menyebabkan kandungan N yang lebih tinggi di IC dibandingkan di SC triticale, baik sebagai persentase maupun sebagai kg ha −1 , yang mengakibatkan interaksi yang signifikan dengan perlakuan pemupukan.
Pada tahun 2010, persentase N yang umumnya lebih tinggi pada lupin tercermin dalam serapan N yang lebih besar pada IC dibandingkan dengan triticale SC (Gambar 4 ), meskipun PLER DM lupin yang disebutkan sebelumnya rendah pada IC. Konsisten dengan pengamatan DM, superioritas IC atas TSC dalam persentase dan serapan N paling menonjol pada perlakuan pemupukan 0 dan P, terutama pada tahap antesis dan adonan susu. Persentase N kedua spesies, baik pada SC atau intercrop pada tahun 2010, juga dianalisis (Tabel 2 ). Rata-rata, tanaman lupin umumnya mencapai persentase N yang lebih tinggi ketika ditanam sebagai SC, dengan pengecualian tahap adonan susu, di mana lupin intercropped memiliki persentase N yang lebih tinggi daripada lupin single-cropped (2,7% vs. 2,5%, p < .05). Tanaman triticale menunjukkan persentase N yang lebih tinggi pada IC dibandingkan dengan SC.
| Bunga pertama Lupin | Bunga Triticale Anthesis | Kematangan adonan susu | Bulir | Jerami | ||
|---|---|---|---|---|---|---|
| Sistem tanam | *** A | *** | *** | *** | *** | |
| LIC | 2.55 | 2.32 | 2.67 | 5.84 | 0.47 | |
| LSC | 2.94 | 2.75 | 2.49 | 5.58 | 0,55 | |
| TIKTAK | 1.63 | 1.20 | 1.05 | 2.41 | 0.34 | |
| TSC | 1.31 | 1.06 | 0,91 | 2.23 | 0,35 | |
| LSD 0,05 | 0.27 | 0.16 | 0.11 | 0.29 | 0,07 | |
| CS × Pemupukan | *** | *** | ** | tidak ada | tidak ada | |
| N | LIC | 2.27 | 2.20 | 2.36 | 5.86 | 0.43 |
| LSC | 3.16 | 2.83 | 2.53 | 5.54 | 0.52 | |
| TIKTAK | 1.67 | 1.25 | 1.08 | 2.39 | 0.34 | |
| TSC | 1.53 | 1.10 | 0,91 | 2.21 | 0.32 | |
| Tidak ada | LIC | 2.02 | 2.21 | 2.66 | 6.06 | 0.48 |
| LSC | 2.71 | 2.20 | 2.13 | 5.65 | 0,55 | |
| TIKTAK | 1.97 | 1.35 | 1.15 | 2.51 | 0.34 | |
| TSC | 1.43 | 1.17 | 1.10 | 2.35 | 0.32 | |
| P | LIC | 2,99 | 2.17 | 2.84 | 5.59 | 0,50 |
| LSC | 2.92 | 3.13 | 2.62 | 5.52 | 0,59 | |
| TIKTAK | 1.55 | 1.11 | 0,94 | 2.39 | 0.34 | |
| TSC | 1.16 | 1.00 | 0.84 | 2.17 | 0.36 | |
| angka 0 | LIC | 2.91 | 2.70 | 2.81 | 5.84 | 0.48 |
| LSC | 2.98 | 2.83 | 2.67 | 5.61 | 0,55 | |
| TIKTAK | 1.31 | 1.10 | 1.01 | 2.35 | 0.34 | |
| TSC | 1.11 | 0,96 | 0,78 | 2.19 | 0.41 | |
| LSD 0,05 | 0.47 | 0.34 | 0.30 | 0,35 | 0.11 |
Singkatan: LSD, perbedaan signifikan terkecil; ns, tidak signifikan. a Hasil Uji F Analisis Varians (ANOVA). * Signifikan untuk p ≤ .05; ** Signifikan untuk p ≤ .01; *** Signifikan untuk p ≤ .001.
Pada semua pengambilan sampel, tidak terdeteksi adanya perbedaan nyata dalam persentase N biomassa triticale antara tanaman yang tumbuh di SC dengan suplemen N dan tanaman yang tumbuh di IC tanpa pemupukan.
Hasil N lupin (kg N ha −1 ) jelas berkurang dengan kehadiran triticale, seperti yang ditunjukkan oleh PLER-nya, yang berkisar dari 0,24 hingga 0,46 (Gambar 5 ). Penurunan ini merupakan konsekuensi dari PLER lupin yang rendah untuk akumulasi DM. PLER rata-rata triticale menurun dari 1,23 pada pengambilan sampel pertama menjadi 0,95 pada saat dewasa, yang menunjukkan bahwa efek positif tumpang sari pada penyerapan N triticale paling menonjol pada tahap awal siklus pertumbuhannya. Perlakuan pemupukan 0 dan P menunjukkan LER tertinggi untuk sifat ini, kemungkinan karena LER yang lebih besar untuk total DM dalam perlakuan ini.

3.5 Panen: Hasil panen dan komponen hasil panen
Hasil gabah bervariasi sesuai musim sebagai respons terhadap variasi gabah/benih m −2 dan menunjukkan interaksi sistem tanam x pemupukan yang dimediasi oleh variasi total DM, indeks panen (HI), bulir per meter persegi, dan polong subur per meter persegi.
Pada tahun 2010, hasil gabah 23% lebih rendah pada LSC dan 44% lebih rendah pada TSC dibandingkan dengan tahun 2012 (Tabel 3 ).
| Jumlah DM tahun 2010 (t ha −1 ) | tahun 2010 Hai | Hasil panen gabah tahun 2010 (t ha −1 ) | Jerami 2010 (t ha −1 ) | Jumlah DM 2012 (t ha −1 ) | tahun 2012 halo | Hasil panen gabah tahun 2012 (t ha −1 ) | Jerami 2012 (t ha −1 ) | |
|---|---|---|---|---|---|---|---|---|
| Sistem tanam | *** A | ** | ** | *** | *** | * | *** | *** |
| IC | 12.8 | 0.32 | 4.1 | 8.7 | 20.1 | 0.34 | 7.9 | 12.2 |
| LSC | 8.6 | 0.38 | 3.3 | 5.4 | 10.3 | 0.41 | 4.2 | 6.0 |
| TSC | 9.9 | 0.42 | 4.1 | 5.8 | 19.4 | 0.38 | 7.3 | 12.1 |
| LSD 0,05 | 0,96 | 0,05 | 0.32 | 1.05 | 2.17 | 0,05 | 0.61 | 1.66 |
| CS × pemupukan | tidak ada | tidak ada | *** | tidak ada | tidak ada | tidak ada | tidak ada | tidak ada |
Singkatan: IC, tumpang sari; lSC, lupin pada tanaman tunggal; LSD, perbedaan signifikan terkecil; ns, tidak signifikan; TSC, triticale pada tanaman tunggal. a Hasil uji F ANOVA. * Signifikan untuk p ≤ .05; ** Signifikan untuk p ≤ .01; *** Signifikan untuk p ≤ .001.
Analisis komponen hasil triticale (Tabel S5 ) mengungkapkan bahwa jumlah gabah per meter persegi yang lebih rendah (15.147 pada tahun 2010 vs 17.978 pada tahun 2012) bertanggung jawab atas hasil gabah triticale yang lebih rendah pada tahun 2010, karena berat gabah sebenarnya lebih tinggi pada tahun 2010 daripada tahun berikutnya (48,7 vs. 42,8 mg). Jumlah gabah yang berkurang pada tahun 2010 terutama karena jumlah bulir fertil per meter persegi yang lebih rendah (267 vs. 303 pada tahun 2012), yang dikaitkan dengan insiden batang steril yang lebih tinggi (25,6 vs. 2,4 batang steril m −2 ). Demikian pula, hasil gabah lupin yang lebih rendah pada tahun 2010 dikaitkan dengan lebih sedikit biji per meter persegi, karena jumlah polong per meter persegi dan biji per polong yang berkurang, meskipun berat biji lebih tinggi pada tahun 2010.
Pada tahun 2012, rata-rata hasil gabah IC lebih tinggi dibanding hasil gabah TSC (Tabel 3 ) karena LER yang sedikit lebih tinggi untuk hasil gabah dan jerami, yang terutama terkait dengan kinerja triticale yang baik, meskipun kontribusi lupin dapat diabaikan, seperti ditunjukkan oleh PLER-nya yang sangat rendah (Gambar 6 ).
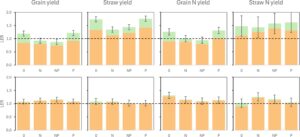
Pada tahun 2010, sebaliknya, rata-rata hasil gabah IC tidak berbeda, tetapi kontribusi lupin, meskipun rendah, menghasilkan interaksi positif untuk hasil gabah dalam IC pada perlakuan 0 (LER = 1,19) dan P (LER = 1,22), satu-satunya perlakuan pemupukan dengan LER lebih besar dari 1. Hasilnya, hasil gabah yang lebih besar tercatat dalam IC dibandingkan dengan TSC hanya pada perlakuan 0 (3,9 vs. 3,2 t ha −1 , p ≤ 0,05) dan P (4,3 vs. 3,6 t ha −1 , p ≤ 0,05), yang dihasilkan dari kombinasi DM yang lebih tinggi (masing-masing sebesar 56% dan 61% pada 0 dan P) dan HI yang lebih rendah (sebesar 20% pada kedua perlakuan) dibandingkan dengan TSC.
Berat biji Triticale tidak terpengaruh oleh penanaman tumpang sari (Tabel S5 ), sedangkan jumlah bulir per meter persegi lebih tinggi dari yang diharapkan yaitu 50% (71% dan 76% pada tahun 2010 dan 2012) karena daya saing lupin yang rendah di IC, yang mengakibatkan jumlah polong fertil per meter persegi yang lebih rendah pada spesies ini, terkait dengan berat biji yang tidak terpengaruh (2012) atau bahkan lebih tinggi (2010, 352 vs. 268 mg, p ≤ .05) dibandingkan di SC.
Untuk menilai apakah lupin dalam IC dapat menggantikan pemupukan N dalam TSC, kami membandingkan biomassa, gabah, dan hasil jerami IC pada perlakuan 0 dengan TSC pada pemupukan N. IC pada 0 menghasilkan total DM yang hampir sama dengan TSC pada N (masing-masing 11,5 dan 11,6 t ha −1 ), tetapi dengan hasil gabah 15% lebih rendah (3,9 vs. 4,6 t ha −1 ) dan hasil jerami 9% lebih tinggi (7,5 vs. 6,9 t ha −1 ).
3.6 Nitrogen saat panen
Pada tahun 2012, keberadaan lupin yang langka di IC secara umum tidak cukup untuk menyebabkan perbedaan dalam hasil atau persentase N antara IC dan TSC. Sebaliknya, pada tahun 2010, persentase N yang lebih tinggi yang menjadi ciri tanaman polong-polongan tercermin dalam persentase N yang lebih tinggi pada biji-bijian IC dibandingkan dengan biji-bijian TSC, serta hasil N yang lebih tinggi (kg N ha −1 ), baik pada biji-bijian maupun jerami (Tabel 4 ).
| Tahun 2010 | Tahun 2012 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N dalam biji-bijian | N dalam jerami | N totalnya DM | N dalam biji-bijian | N dalam jerami | N totalnya DM | ||||||||
| (%) | (kg ha −1 ) | (%) | (kg ha −1 ) | (%) | (kg ha −1 ) | (%) | (kg ha −1 ) | (%) | (kg ha −1 ) | (%) | (kg ha −1 ) | ||
| Sistem tanam | *** A | *** | ** | ** | *** | *** | *** | *** | *** | ** | *** | *** | |
| IC | 3.05 | 125 | 0.37 | 32.1 | 1.24 | 157 | 1.34 | 119 | 0.34 | 43.2 | 0,80 | 162 | |
| LSC | 5.58 | 182 | 0,55 | 30.1 | 2.48 | 212 | 4.97 | 210 | 0,55 | 39.2 | 2.37 | 243 | |
| TSC | 2.23 | 92 | 0.36 | 19.9 | 1.15 | 112 | 1.39 | 102 | 0.32 | 33.1 | 0.73 | 141 | |
| LSD 0,05 | 0.31 | 1.8 | 0,08 | 0.7 | 0.27 | 2.1 | 0.51 | 2.8 | 0,07 | 0.4 | 0.29 | 2.8 | |
| CS × Pemupukan | tidak ada | * | tidak ada | tidak ada | tidak ada | tidak ada | * | *** | tidak ada | tidak ada | * | *** | |
| angka 0 | IC | 3.45 | 136 | 0.38 | 29.0 | 1.45 | 165 | 1.11 | 108 | 0.29 | 32.2 | 0.73 | 140 |
| LSC | 5.61 | 203 | 0,55 | 29.0 | 2.69 | 232 | 5.21 | 191 | 0.57 | 28.3 | 2.53 | 219 | |
| TSC | 2.20 | 70 | 0.41 | 17.0 | 1.20 | 87 | 1.18 | 82 | 0.29 | 32.4 | 0.63 | 115 | |
| N | IC | 2.78 | 114 | 0.36 | 31.9 | 1.11 | 146 | 1.40 | 136 | 0.38 | 56.5 | 0,91 | 193 |
| LSC | 5.54 | 186 | 0.52 | 29.9 | 2.42 | 215 | 4.60 | 195 | 0,50 | 29.6 | 2.20 | 225 | |
| TSC | 2.22 | 103 | 0.31 | 21.7 | 1.08 | 125 | 1.52 | 120 | 0.36 | 46.2 | 0,80 | 166 | |
| Tidak ada | IC | 2.91 | 119 | 0.36 | 33.3 | 1.15 | 152 | 1.60 | 128 | 0.39 | 45.1 | 0,88 | 173 |
| LSC | 5.65 | 169 | 0,55 | 31.7 | 2.32 | 200 | 4.93 | 188 | 0,55 | 31.2 | 2.33 | 219 | |
| TSC | 2.36 | 116 | 0.32 | 21.0 | 1.20 | 137 | 1.64 | 117 | 0.36 | 40.4 | 0.86 | 158 | |
| P | IC | 3.05 | 130 | 0.37 | 34.2 | 1.24 | 164 | 1.23 | 103 | 0.30 | 39.1 | 0,70 | 142 |
| LSC | 5.52 | 172 | 0,59 | 29.9 | 2.50 | 202 | 5.14 | 267 | 0.57 | 43.3 | 2.41 | 310 | |
| TSC | 2.15 | 78 | 0.38 | 20.2 | 1.11 | 98 | 1.22 | 88 | 0.29 | 37.7 | 0.62 | 126 | |
| LSD 0,05 | 0.44 | 3.0 | 0,08 | 1.0 | 0,25 | 3.8 | 0.47 | 2.9 | 0.14 | 1.4 | 0.21 | 3.6 | |
Catatan : Perlakuan pemupukan: 0, tanpa pemupukan; N, 100 kg ha -1 N; P, 39 kg ha -1 P; NP, 100 kg ha -1 N + 39 kg ha -1 P. Singkatan: DM, bahan kering; IC, tanaman sela; LSC, lupin pada tanaman tunggal; LSD, perbedaan signifikan terkecil; ns, tidak signifikan; TSC, triticale pada tanaman tunggal. a Hasil uji F ANOVA. * Signifikan untuk p ≤ .05; ** Signifikan untuk p ≤ .01; *** Signifikan untuk p ≤ .001.
Konsisten dengan data hasil gabah, pada tahun 2010, perlakuan pemupukan 0 dan P merupakan satu-satunya yang memiliki LER lebih besar dari 1 untuk hasil gabah N, karena PLER lupin yang lebih tinggi (Gambar 6 ), dan ini menyebabkan IC mengungguli TSC hanya pada perlakuan 0 (136 kg N ha −1 pada IC vs. 70 kg N ha −1 pada TSC) dan P (130 kg N ha −1 pada IC vs. 78 kg N ha −1 pada TSC). Hal ini kemungkinan merupakan hasil dari keunggulan perlakuan pemupukan ini dalam produksi DM.
Total hasil N pada IC pada perlakuan pemupukan 0 adalah 165 kg ha −1 pada saat matang, yaitu 40 kg N ha −1 lebih besar dari total hasil N triticale di SC yang menerima 100 kg N ha −1 pupuk.
4 DISKUSI
Penelitian sebelumnya telah menunjukkan peningkatan hasil dan kualitas pada lupin yang ditanam secara tumpangsari dengan berbagai spesies serealia, seperti oat (McKenzie & Spaner, 1999 ), jagung (Carruthers et al., 2000 ), dan barley (Strydhorst et al., 2008 ). Akan tetapi, hanya sedikit penelitian yang menganalisis tanaman sela lupin–triticale dalam uji lapangan, dan penelitian tersebut dilakukan di iklim dengan musim dingin yang mengharuskan penanaman di musim semi (Azo et al., 2012 ; Gałęzewski, 2020 ; Górski & Płaza, 2024 ; Księżak & Staniak, 2013 ). Hanya Carton et al. ( 2020 ) yang merujuk pada tanaman sela triticale–lupin yang ditanam pada musim gugur di Prancis, tetapi dalam konteks penekanan gulma.
Meskipun daya saing lupin rendah ketika ditanam dengan triticale pada tingkat penanaman yang digunakan di sini, data kami tahun 2012 menunjukkan bahwa tanaman sela triticale–lupin dapat sama produktifnya dengan TSC, berkat anakan triticale yang melimpah, bahkan ketika lupin hampir sepenuhnya ditekan oleh triticale. Di sisi lain, data tahun 2010 memberikan wawasan berharga tentang komplementaritas antara kedua spesies ini karena lupin mampu memberikan kontribusinya pada tanaman sela berkat daya saing triticale yang lebih rendah, yang berasal dari tekanan genangan air musim ini yang berasal dari kombinasi curah hujan musim dingin yang melimpah dan tekstur tanah liat, yang sering terjadi di musim dingin dalam jenis lingkungan ini (Bassu et al., 2009 ).
Berdasarkan hipotesis bahwa lupin dapat ditanam secara tumpang sari dengan triticale dengan penanaman musim gugur dan tanpa irigasi di lingkungan Mediterania, kami melaksanakan penelitian eksplorasi di alam, menggunakan pengambilan sampel DM dan N berulang dan pengukuran intersepsi radiasi untuk menghubungkan keuntungan terkait N yang diketahui dari penanaman sela serealia-kacang-kacangan dengan perolehan radiasi dan efisiensi konversi radiasi menjadi DM.
4.1 Intersepsi radiasi dan pertumbuhan lupin di IC terhambat oleh perawakannya yang lebih pendek dan laju perkembangan yang lebih lambat
Dengan meneliti dinamika intersepsi dan pertumbuhan radiasi (akumulasi DM), kami memperoleh wawasan tentang mekanisme yang mendorong persaingan asimetris antara triticale dan lupin. Dalam percobaan kami, perkembangan luas daun lupin yang lebih lambat, perawakan yang lebih pendek, dan kebiasaan semi-determinat merugikannya dalam tumpang sari, yang menyebabkan berkurangnya IPAR.
Ukuran benih dan bibit dapat memengaruhi laju pertumbuhan awal dan kemampuan bersaing (Andersen et al., 2007 ), tetapi data evolusi FIPAR kami menunjukkan bahwa laju perkembangan juga memainkan peran penting dalam membentuk interaksi antarspesies dalam intercropping sereal–kacang-kacangan. Suhu dasar lupin yang lebih tinggi (3,0–4,5°C, Duthion et al., 1994 ) dibandingkan dengan triticale (0°C) (Giunta et al., 2001 ) berkontribusi pada perkembangan luas daunnya yang lebih lambat, yang memberikan triticale keuntungan kompetitif awal yang biasanya meningkat sepanjang musim. Ini, dikombinasikan dengan RUE legum yang lebih rendah dibandingkan dengan sereal (Hay & Porter, 2006 ), menyebabkan PLER rendah untuk lupin dalam hal total DM, yang secara progresif mengurangi komplementaritas dengan triticale. Dominasi kompetitif sereal atas legum telah diamati pada intercropping lain, seperti kacang polong–gandum (van Kessel & Hartley, 2000 ).
Perawakan lupin yang lebih pendek juga berdampak negatif pada kinerjanya ketika ditanam secara tumpang sari dengan jagung (Carruthers et al., 2000 ) dan barley (Strydhorst et al., 2008 ). Dalam penelitian kami, tumpang tindih pertumbuhan biji dan polong lupin dengan pembentukan booting dan pemanjangan tangkai triticale, terutama pada tahun 2012 ketika triticale tidak terpengaruh oleh genangan air, mengakibatkan hasil gabah lupin yang lebih rendah. Oceania, kultivar triticale yang digunakan, dipilih karena potensi hasil yang tinggi dan perawakannya yang rendah, tetapi kebiasaan pertumbuhan awal musim semi berkontribusi pada penalti lupin. Kultivar triticale musim semi dicirikan oleh perkembangan yang lebih cepat daripada kultivar triticale musim dingin dan karena itu berpotensi lebih kompetitif terhadap spesies yang bertubuh lebih pendek dan berkembang lambat seperti lupin. Pengurangan hasil lupin berpotensi dikurangi dengan menggunakan kultivar musim dingin alih-alih kultivar musim semi. Alternatifnya, penanaman sela alih-alih penanaman sela baris (Gałęzewski, 2020 ) atau penanaman lupin lebih awal dapat dipertimbangkan, meskipun hal ini menghadirkan tantangan teknis di tanah liat selama bulan-bulan hujan.
4.2 Keunggulan tanaman sela triticale–lupin dibandingkan tanaman tunggal triticale bergantung pada pemupukan.
Komplementaritas antara triticale dan lupin untuk DM dan hasil N dipengaruhi oleh perlakuan pemupukan, dengan perlakuan pemupukan 0 dan P umumnya menghasilkan LER tertinggi untuk kedua sifat tersebut. LER mencapai nilai setinggi 1,7 untuk DM pada antesis triticale, didorong oleh PLER triticale yang tinggi. Ini adalah hasil yang menarik karena tahap polong hijau pipih dari lupin, yang secara kasar sesuai dengan tahap antesis triticale, adalah tahap di mana hasil DM dimaksimalkan dalam tumpang sari kacang polong–triticale 50:50 (Płaza & Górski, 2024 ), dan kandungan lemak kasar dimaksimalkan dalam tumpang sari triticale–lupin (Górski & Płaza, 2023 ).
Hasil kami mengonfirmasi potensi lebih besar yang biasanya diekspresikan oleh penanaman tumpang sari legum/sereal dalam sistem dengan ketersediaan N rendah, di mana keseimbangan antara fiksasi N dan penyerapan N tanah oleh kedua spesies memungkinkan penyesuaian ketersediaan N untuk memenuhi kebutuhan sereal (Carton et al., 2020 ; Demie et al., 2022 ; Hauggaard-Nielsen et al., 2003 ; Naudin et al., 2010 ). Dapat dihipotesiskan bahwa komplementaritas untuk produksi DM dalam percobaan ini sebagian berasal dari RUE IC yang lebih tinggi dibandingkan dengan TSC di bawah perlakuan pemupukan 0, kemungkinan terkait dengan persentase N DM IC yang lebih tinggi (Sinclair & Horie, 1989 ). RUE yang lebih tinggi juga telah dilaporkan dalam campuran jagung dan sorgum dengan legum dibandingkan dengan SC (Umesh et al., 2023 ). Hasil baik yang diamati dengan perlakuan P, di sisi lain, dapat dikaitkan dengan peran utama fosfor dalam fiksasi N simbiosis lupin (Zhong et al., 2023 ).
Dalam tanaman sela lupin–sereal, komplementaritas juga dapat muncul dari morfologi sistem perakaran yang berbeda, dengan akar sereal yang lebih halus dan lebih dangkal menjelajahi lapisan tanah atas, yang memaksa akar tunggang lupin (Kemper et al., 2022 ; Mariotti et al., 2009 ; Wiche et al., 2016 ) untuk mengambil nutrisi dan air dari lapisan yang lebih dalam (Górski & Płaza, 2024 ). Pengaruh tingkat perkembangan yang berbeda dari kedua spesies pada ekspresi komplementaritas juga dapat dihipotesiskan: sementara triticale menghasilkan lebih banyak biomassa daripada lupin pada tahap pertumbuhan awal, laju pertumbuhannya menurun setelah antesis, bertepatan dengan peningkatan pertumbuhan lupin, yang memungkinkan komplementaritas dalam penggunaan sumber daya dari waktu ke waktu, seperti yang dicatat oleh Carton et al. ( 2020 ).
Hasil pada total DM menyoroti bahwa, tergantung pada tahap pertumbuhan dan tanpa pemupukan, 49%–71% lebih banyak luas permukaan akan diperlukan untuk mencapai hasil DM yang sama dengan triticale SC seperti yang diperoleh dengan intercrop triticale–lupin. LER yang lebih rendah—berkisar dari 1,07 hingga 1,37, tergantung pada proporsi kedua spesies—dilaporkan untuk intercrop triticale–lupin yang ditanam tanpa pemupukan di Polandia (Górski & Płaza, 2024 ) dan untuk intercrop gandum–kacang polong (Ghaley et al., 2005 ). Berbeda dengan temuan kami, hasil DM dari intercrop gandum–kacang polong tanpa pemupukan N (370 ± 28 g m −2 ) yang dicatat oleh Ghaley et al. ( 2005 ) lebih rendah dibandingkan dengan hasil DM (478 ± 47 g m −2 ) gandum di SC dengan pemupukan N 8 g m −2 .
IC lebih unggul daripada TSC dalam hal persentase N total DM, tidak hanya karena adanya sejumlah kecil DM lupin dengan persentase N yang secara inheren tinggi, tetapi juga karena tanaman triticale yang tumbuh di IC memiliki persentase N yang lebih tinggi daripada yang ada di SC. Sinkronisasi yang lebih besar antara dinamika temporal kebutuhan N serealia dan ketersediaan N tanah mungkin telah berkontribusi pada persentase N yang lebih tinggi dari triticale yang ditanam di sela-sela (Stomph et al., 2020 ). Azo et al. ( 2012 ) juga mengamati peningkatan nilai nutrisi hijauan pada sela-sela triticale–lupin, yang tidak hanya terkait dengan persentase N total DM dan persentase protein kasar yang lebih tinggi tetapi juga kandungan serat yang lebih tinggi. Keuntungan sela dalam hal persentase N kurang dari yang diharapkan mengingat persentase N yang tinggi dari LSC (2,9% pada pengambilan sampel pertama menjadi 2,7% pada pengambilan sampel kedua), kemungkinan karena rendahnya proporsi DM lupin pada sela-sela.
Efek gabungan pada DM dan N% memungkinkan intercrop untuk menunjukkan komplementaritas untuk total hasil N juga. Keuntungan terbesar IC atas TSC dalam total hasil N diamati pada tahap adonan susu (+65%) dan pada saat matang (+40%). Ini kontras dengan intercrop triticale–kacang polong yang ditanam di Polandia oleh Płaza dan Górski ( 2024 ), di mana tahap antesis triticale (hampir bersamaan dengan tahap polong hijau pipih lupin) memberikan hasil DM tertinggi tetapi hasil protein sedikit lebih rendah daripada tahap pembungaan kacang polong sebelumnya. Peningkatan yang diamati dalam total hasil N sejalan dengan peningkatan 50% yang dilaporkan dalam intercrop gandum durum–kacang arab (Latati et al., 2019 ), dan melebihi +27% yang diamati dalam intercrop barley–lupin (Strydhorst et al., 2008 ) dibandingkan dengan SC sereal masing-masing. Sekali lagi, komplementaritas ditingkatkan dengan tidak adanya pemupukan N, konsisten dengan temuan Jensen ( 1996 ) dalam percobaan kacang polong–barley. Komplementaritas penggunaan N yang tinggi dapat dicapai ketika tidak ada pupuk yang diberikan, karena serealia mengambil bagian N anorganik tanah yang tidak proporsional, yang memaksa legum untuk memenuhi kebutuhan N-nya melalui fiksasi simbiosis.
Seperti yang diamati dalam Carton et al. ( 2020 ), persaingan dari triticale mengurangi hasil gabah lupin, kemungkinan karena ketidakmampuan lupin untuk mencapai intersepsi cahaya maksimum segera setelah berbunga, yang merupakan periode kritis untuk pembentukan polong dan pengisian biji lupin (Cowling et al., 1998 ), karena perkembangan luas daun triticale yang lebih cepat. Namun, IC menghasilkan hasil gabah yang lebih tinggi daripada kedua SC di bawah perlakuan 0 dan P pada tahun 2010, karena DM yang lebih besar yang dibahas sebelumnya, yang berarti IC bahkan lebih menguntungkan untuk hasil jerami daripada untuk hasil gabah. Nilai LER sebesar 1,20 yang tercatat untuk hasil gabah lebih tinggi daripada 1,14 yang diamati dalam intercrop barley-kacang polong (Cowden et al., 2020 ), tetapi lebih rendah dari 1,38 yang dilaporkan dalam studi intercrop lentil-gandum (Cibella et al., 1995 ). Mirip dengan studi-studi ini, pemupukan berdampak negatif pada komplementaritas untuk hasil gabah.
Selain itu, persentase N biji-bijian mendapat manfaat dari tumpang sari, khususnya tanpa adanya masukan N, sesuai dengan sebagian besar literatur tentang tumpang sari serealia-kacang-kacangan (Bedoussac & Justes, 2010 ; Gooding et al., 2007 ; Jensen, 1996 ; Knudsen et al., 2004 ; Naudin et al., 2010 ). Seperti yang dibahas sehubungan dengan DM, N% biji-bijian IC yang lebih tinggi tidak hanya karena adanya biji lupin dalam biji-bijian yang dipanen, tetapi juga karena N% yang lebih besar dalam biji-bijian triticale yang ditanam tumpang sari dengan lupin dibandingkan dengan yang ditanam di SC. Tumpang sari serealia dengan kacang-kacangan telah terbukti secara umum meningkatkan konsentrasi protein biji-bijian serealia dibandingkan dengan penanaman tunggal, khususnya dalam kondisi N tanah rendah dan masukan N rendah (Księżak et al., 2023 ; Stomph et al., 2020 , dan referensi di dalamnya). Yang perlu diperhatikan, pada IC yang tidak diberi pupuk, konsentrasi N pada biji triticale sebanding dengan triticale yang ditanam tunggal dan diberi pupuk N sebanyak 100 kg ha −1 (perlakuan N), seperti yang sebelumnya dilaporkan oleh Naudin et al. ( 2010 ) pada tanaman sela gandum/kacang polong.
5 KESIMPULAN
Studi ini, yang pertama kali menganalisis tumpang sari triticale–lupin di lingkungan Mediterania, menunjukkan bahwa meskipun terdapat perbedaan musim yang nyata dan pengaruhnya terhadap proporsi bahan kering lupin dalam tumpang sari, produktivitas tumpang sari triticale–lupin sebanding dengan—atau bahkan lebih tinggi daripada—penanaman tunggal triticale.
Komplementaritas antara kedua spesies tersebut terbukti untuk DM dan hasil N, sejalan dengan penelitian sebelumnya pada tanaman sela serealia-kacang-kacangan lainnya, tetapi penggunaan dua spesies ‘yatim piatu’, lupin dan triticale, lebih lanjut mendorong diversifikasi tanaman. Komplementaritas yang diamati tanpa adanya pupuk menunjukkan bahwa tanaman sela ini dapat sangat cocok untuk sistem pertanian input rendah di lingkungan Mediterania. Selain itu, penanaman sela triticale dengan lupin telah terbukti secara efektif menggantikan pemupukan N tanpa mengorbankan DM atau kandungan N-nya dalam pakan ternak.
Pendekatan eksperimental memungkinkan hubungan manfaat terkait N dalam tanaman sela serealia-kacang-kacangan dengan perolehan radiasi dan efisiensi konversi menjadi bahan baku. Pendekatan ini juga menyoroti peran penting laju perkembangan, khususnya sinkronisasi perkembangan tajuk, yang penting untuk menyeimbangkan persaingan antar spesies ketika perbedaan ketinggian signifikan. Oleh karena itu, pemilihan kultivar dalam spesies yang ditanam sela sangat penting bagi keberhasilan sistem serealia-kacang-kacangan. Secara khusus, penggunaan kultivar triticale musim dingin untuk penanaman sela dengan lupin sebagai pengganti triticale musim semi direkomendasikan untuk mencapai kontribusi yang lebih seimbang antara kedua spesies.



Leave a Reply