
Abstrak
LATAR BELAKANG
Cyantraniliprole, insektisida diamida, digunakan secara luas di Brasil untuk mengendalikan hama penghisap dan penggugur daun, termasuk ulat grayak ( Spodoptera frugiperda ), hama pertanian utama. Namun, meningkatnya resistensi terhadap diamida telah menimbulkan kekhawatiran tentang efektivitas jangka panjang cyantraniliprole. Penelitian ini bertujuan untuk (i) menilai evolusi resistensi cyantraniliprole pada populasi lapangan S. frugiperda , (ii) menyelidiki resistensi silang terhadap diamida lain, dan (iii) menganalisis peran mutasi I4790K dalam mekanisme resistensi.
HASIL
Penurunan signifikan dalam kerentanan terhadap siantraniliprole diamati pada populasi lapangan S. frugiperda di Brasil, khususnya di wilayah dengan sistem pertanian intensif, seperti Cerrado Brasil, dari tahun 2017 hingga 2023. Galur S. frugiperda yang resistan terhadap siantraniliprole dipilih dari populasi yang dikumpulkan di lapangan di Bahia (BA) menggunakan metode penyaringan F 2 . Pewarisan resistensi terhadap siantraniliprole pada galur ini bersifat resesif autosomal dan monogenik, dengan rasio resistensi 3414 kali lipat. Resistensi silang yang tinggi terhadap diamides flubendiamide, chlorantraniliprole, dan cyclaniliprole juga terdeteksi. Analisis molekuler mengonfirmasi keberadaan mutasi I4790K homozigot pada gen reseptor ryanodine sebagai salah satu mekanisme resistensi galur yang resistan terhadap siantraniliprole ini.
KESIMPULAN
Temuan ini menyoroti penurunan kerentanan populasi lapangan S. frugiperda terhadap cyantraniliprole dan peran mutasi I4790K dalam mempercepat evolusi resistensi terhadap insektisida diamida karena resistensi silang. Hasil ini menggarisbawahi kebutuhan mendesak untuk strategi pengelolaan hama terpadu (IPM), termasuk rotasi insektisida dan pemantauan resistensi, untuk menjaga kemanjuran cyantraniliprole dan insektisida lainnya. © 2025 Penulis. Ilmu Pengelolaan Hama diterbitkan oleh John Wiley & Sons Ltd atas nama Society of Chemical Industry.
1. PENDAHULUAN
Produksi pangan global diproyeksikan meningkat sebesar 36–56% pada tahun 2050, 1 , 2 dan hama serangga merupakan tantangan yang signifikan dalam mencapai tujuan ini. Pengelolaan hama ini terutama bergantung pada insektisida sintetis, khususnya tanaman seperti jagung dan kapas, dan tanaman yang dimodifikasi secara genetika yang mengekspresikan protein Cry dari Bacillus thuringiensis (Bt). Namun, penggunaan insektisida yang berlebihan telah menimbulkan kekhawatiran mengenai dampaknya terhadap lingkungan dan kesehatan manusia. Oleh karena itu, berbagai upaya sedang dilakukan untuk mengembangkan insektisida selektif yang mempertahankan kemanjuran sekaligus meningkatkan keselamatan lingkungan dan manusia. 3
Di antara insektisida yang lebih baru, diamida menonjol karena tindakannya yang selektif, toksisitas rendah terhadap organisme non-target, dan profil lingkungan yang menguntungkan. 4 Karakteristik ini telah berkontribusi pada pangsa pasar diamida sekitar 8% dari pasar insektisida global dalam beberapa tahun terakhir. 3 Strategi pemasaran utama untuk kelompok senyawa ini adalah mempromosikan cara kerja barunya, yang memberikan pengendalian spektrum luas terhadap berbagai hama, termasuk serangga penghisap dan penggugur daun. 5 – 7
Diamida juga digunakan secara luas sebagai perlakuan benih karena translokasi tanaman dan aktivitas residunya yang tinggi, sehingga memberikan perlindungan awal yang penting bagi tanaman saat tanaman tersebut rentan terhadap serangan hama. 8 , 9 Dua diamida pertama, flubendiamida dan klorantraniliprole, didaftarkan pada tahun 2007 untuk mengendalikan hama lepidoptera. 6 Pada tahun 2013, molekul baru seperti siantraniliprole diperkenalkan, yang memperluas spektrum pengendalian sekaligus mempertahankan selektivitas untuk spesies non-target. 7 Siantraniliprole sangat efektif terhadap serangga pemakan getah, termasuk lalat putih, wereng, kutu daun, psyllid, dan lepidoptera, sehingga menjadikannya alat yang berharga untuk program pengelolaan hama terpadu. 4 , 7 , 10
Di Brasil, sistem pertanian dicirikan oleh lahan subur yang luas dan kondisi iklim yang mendukung, yang berkontribusi terhadap peningkatan populasi hama secara signifikan. 11 Sistem ini, terutama pada tanaman kedelai, jagung, dan kapas, dapat bertahan hingga tiga siklus penanaman tahunan di beberapa wilayah, khususnya di Cerrado Brasil, yang memiliki lahan subur terbesar di negara bagian Bahia (BA), Mato Grosso (MT), Goiás (GO), Mato Grosso do Sul (MS), dan Minas Gerais (MG). 12 Di Brasil, wilayah penghasil biji-bijian biasanya memiliki tiga musim panen yang berbeda: musim pertama (Oktober hingga Januari), musim kedua (Februari hingga Mei, yang menandai berakhirnya musim hujan), dan musim ketiga/di luar musim (Juni hingga September). Lingkungan ini khususnya menguntungkan hama polifag seperti ulat grayak, Spodoptera frugiperda (JE Smith, 1797) (Lepidoptera: Noctuidae), hama yang penting secara global dan salah satu hama utama tanaman jagung, yang juga semakin banyak dilaporkan pada tanaman kapas dan kedelai di Brasil. 13 – 15
Paparan terus-menerus S. frugiperda terhadap insektisida dan protein Bt telah menyebabkan resistensi yang berevolusi di lapangan terhadap beberapa insektisida, termasuk avermectina, 16 benzoilurea, 17 , 18 spinosin, 19 , 20 organofosfat, 21 diamida, 22 , 23 dan protein Bt. 24-28 Resistensi terhadap diamida terutama terjadi melalui mutasi pada gen reseptor ryanodine, yang mengurangi afinitas pengikatan insektisida, yang mengarah pada ketidakpekaan. 29-31 Mekanisme resistensi ini pertama kali diidentifikasi pada Plutella xylostella (Lepidoptera: Plutellidae) pada tahun 2012, dengan mutasi pada daerah C-terminal dari domain transmembran reseptor ryanodine. 32 , 33 Penelitian selanjutnya telah mengonfirmasi bahwa perubahan pada asam amino tertentu, khususnya G4946E, I4790M, dan I4790K, sangat penting untuk memberikan resistensi terhadap diamida. Mutasi I4790K pada P. xylostella telah dikaitkan dengan tingkat resistensi silang yang tinggi di antara semua diamida, yang memengaruhi diamida antranilik dan yang berasal dari asam ftalat. 29 , 34 Laporan terbaru menunjukkan bahwa mutasi I4790M dan I4790K hadir dalam populasi S. frugiperda di Brasil . 23 Namun, peran mutasi I4790K pada individu homozigot masih belum jelas, karena hanya beberapa individu dengan genotipe I/K yang telah didokumentasikan hingga saat ini.
Penelitian ini bertujuan untuk mengevaluasi perubahan kerentanan populasi ulat grayak di ladang terhadap cyantraniliprole selama musim tanam yang berbeda dari tahun 2017 hingga 2023. Lebih jauh, kami bertujuan untuk memilih dan mengkarakterisasi galur S. frugiperda yang resistan terhadap cyantraniliprole untuk memahami pewarisan resistensi dan resistensi silang terhadap insektisida diamida lainnya. Temuan dari penelitian ini akan berkontribusi untuk memahami evolusi S. frugiperda terhadap cyantraniliprole dan menerapkan strategi untuk mengelola resistensi ini.
2 BAHAN DAN METODE
2.1 Serangga
Galur yang rentan terhadap S. frugiperda (SUS) yang digunakan dalam bioassay dibesarkan dengan diet buatan 35 tanpa paparan insektisida atau toksin Bt selama beberapa generasi dalam kondisi laboratorium. Galur yang resistan terhadap insektisida cyantraniliprole (CYA-R) dipilih dari individu yang bertahan hidup menggunakan metode penyaringan F 2 36 dari populasi yang dikumpulkan di Luis Eduardo Magalhães, Bahia, Brasil (11°44′48.90″ S, 45°46′07.60″ W). Setelah seleksi, kurva respons-konsentrasi diperkirakan dalam CYA-R generasi F 3 untuk menentukan konsentrasi tertinggi untuk tekanan seleksi untuk memilih individu homozigot (~10% dari kelangsungan hidup). Dari generasi F 4 , CYA-R dipertahankan pada tekanan seleksi konstan pada konsentrasi cyantraniliprole 3200 μg ai mL −1 .
Bahasa Indonesia: Untuk pemantauan kerentanan S. frugiperda terhadap cyantraniliprole, 1000–1500 larva awalnya dikumpulkan dari populasi lapangan di daerah-daerah penghasil jagung utama di Brasil dari tahun panen 2017 hingga 2023 (Tabel S1 ). Setelah pengumpulan, proses penyaringan dilakukan untuk memastikan viabilitas dan kondisi larva. Setelah triase ini, larva yang layak diangkut ke laboratorium kami, di mana mereka dibesarkan dengan diet buatan hingga mencapai tahap pupa. Setelah disinfeksi dengan larutan tembaga sulfat 6,5%, pupa ditempatkan di kandang PVC silinder (diameter 10 cm × tinggi 20 cm) yang dilapisi kertas putih dan ditutup di bagian atas dengan wadah plastik. Orang dewasa diberi makan larutan madu:air 1:10. Massa telur dikumpulkan setiap 2 hari dan ditempatkan dalam gelas plastik (100 mL) hingga larva menetas. Larva neonatus dipindahkan ke gelas plastik baru yang berisi makanan buatan, di mana mereka tetap berada di sana hingga instar ketiga, saat mereka digunakan dalam bioassay. Semua tahap perkembangan dipelihara di ruangan dengan suhu terkontrol (25 ± 2 °C, kelembaban relatif 60 ± 10%, dan periode cahaya 14:10 jam [L:D]).
2.2 Pemantauan kerentanan Spodoptera frugiperda terhadap cyantraniliprole
Metode bioassay diet-overlay digunakan untuk memantau kerentanan S. frugiperda terhadap cyantraniliprole (Benevia® OD, 100 g ai l −1 , FMC Química do Brasil LTDA, Campinas, SP, Brazil). Bioassay dilakukan pada plat akrilik 24-sel (dengan luas 1,9 cm 2 in) (Costar® Maizeing Incorporated, Sigma-Aldrich Brazil Ltd., São Paulo, BR), yang berisi 1,25 mL diet buatan per sumur, di mana 30 μL larutan insektisida dan surfaktan (Triton® X-100, Labsynth, Diadema, SP, Brazil) pada konsentrasi 0,1% (v/v) dipipet ke setiap sumur. Konsentrasi diagnostik adalah 180 μg ai mL −1 37 dan setelah mengeringkan larutan insektisida, 480 larva instar ketiga diinfestasi dengan setiap populasi lahan (satu larva/sumur), dengan total 20 ulangan dari 24 larva. Plat diinkubasi dalam ruang dengan suhu terkontrol (25 ± 2 °C, kelembaban relatif 60 ± 10%, fotoperiode 14:10 jam [L:D]), dan mortalitas dinilai 96 jam setelah paparan. Larva yang gagal bergerak normal saat disentuh dengan sikat dianggap mati.
Sebuah meta-analisis dilakukan untuk mengevaluasi kelangsungan hidup S. frugiperda yang terpapar cyantraniliprole, menggunakan data kelangsungan hidup yang dikumpulkan selama penelitian kami (2017–2023). Analisis ini juga menggabungkan data yang tersedia dalam literatur mengenai kemanjuran dua insektisida diamida lainnya yang diuji di Brasil. 37 – 39 Untuk meta-analisis ini, data kelangsungan hidup terhadap cyantraniliprole dianalisis menggunakan uji Kruskal–Wallis non-parametrik, 40 diikuti oleh perbandingan rata-rata menggunakan uji Dunn 41 dengan koreksi Bonferroni. Kerentanan populasi lapangan S. frugiperda terhadap cyantraniliprole, chlorantraniliprole, dan flubendiamide dinilai melalui pemantauan data historis dari literatur 37 – 39 dan penelitian kami. Model campuran linier umum (GLMM) dipasang menggunakan Pembuat Model Template, yang diimplementasikan dalam paket ‘glmmTMB’ 42 Model ini mengasumsikan distribusi binomial menggunakan istilah penghalusan dalam rumus model GAM, dengan insektisida dan Negara asal populasi sebagai variabel acak. Analisis grafis dilakukan menggunakan nilai prediksi dari GLMM, menggunakan metode ‘loess’ untuk membuat regresi untuk variabel respons kelangsungan hidup yang diprediksi. Semua analisis dan visualisasi dilakukan dalam perangkat lunak R versi 4.3.1. 43
2.3 Layar F 2 untuk memperkirakan frekuensi alel resistensi
Untuk memperkirakan frekuensi alel resistensi menggunakan metode penyaringan F 2 36 kami mengevaluasi tujuh populasi yang dikumpulkan di wilayah dengan sistem penanaman jagung intensif di Brasil (hingga tiga musim tanam per tahun pertanian) antara tahun tanam 2022 dan 2023 (Tabel 1 ). Pada tahap pupa, individu dipisahkan berdasarkan jenis kelamin, dan sekitar 190 pasangan terbentuk dan dianggap isoline. Setiap pasangan dipelihara dalam gelas plastik 500 mL yang berisi larutan madu:air 1:10, dan massa telur dikumpulkan setiap 2 hari. Setelah keturunan F 1 mencapai instar ketiga, mereka dipindahkan ke piring 32 sel yang berisi diet buatan. Tiga replikasi per isoline (96 larva) dipelihara hingga menjadi pupa.
| Kode | Lokasi (Kota, Negara Bagian) | N † | Tahun Panen ‡ | Tanggal tiba | Lintang (S) | Bujur (W) |
|---|---|---|---|---|---|---|
| PERGI-19 | Kristalina, GO | tahun 1168 | Tahun 2022/1 | 27 Oktober 2021 | 16°29′59.80″ | 47°36′36.80″ |
| BA-9 | Luis Eduardo Magalhães, BA | tahun 1400 | Tahun 2022/1 | 11 Maret 2021 | 11°44′48.90″ | 45°46′07.60″ |
| MT-20 | Lucas dari Rio Verde, MT | tahun 1100 | Tahun 2022/2 | 22 Maret 2022 | 13°01′38.30″ | 55°57′11.40″ |
| Bahasa Indonesia: MS-14 | Chapadão do Sul, MS | 730 | Tahun 2022/2 | 19 April 2022 | 18°44′23.54″ | 52°31′03.02″ |
| C_BA-6 (katun) | Luis Eduardo Magalhães, BA | 894 | Tahun 2022/3 | 31 Mei 2022 | 12°01′41.84″ | 45°45′02.32″ |
| PERGI-22 | Santa Helena, GO | tahun 830 | Tahun 2023/1 | 26 Oktober 2022 | 17°51′37.09″ | 50°23′34.07″ |
| BA-10 | Sao Desiderio, BA | tahun 1226 | Tahun 2023/1 | 14 Desember 2022 | 13°40′15.32″ | 46°07′37.54″ |
† Jumlah serangga yang diuji. ‡ Tahun panen dimulai pada bulan Oktober tahun sebelumnya dan berakhir pada bulan September tahun berikutnya, dibagi menjadi sekitar tiga musim, masing-masing musim berdurasi 4 bulan.
Prosedur yang sama digunakan untuk pengembangbiakan dan pembuatan genotipe generasi F2 . Kandang yang berisi keturunan dewasa F1 dari setiap isolin dipelihara untuk mengumpulkan telur dari generasi F2 untuk pemantauan. Bioassay adalah overlay diet di mana diet diperlakukan dengan cyantraniliprole pada konsentrasi diskriminatif 180 μg ai mL −1 . Untuk setiap isolin, sekitar 120 larva instar ketiga diuji, dan isolin yang menunjukkan kelangsungan hidup lebih besar dari 6,25% dianggap positif, berdasarkan pada premis bahwa 1 dari setiap 16 individu yang berasal dari induk yang rentan dan heterozigot yang dikumpulkan dari lapangan akan menunjukkan homozigositas untuk alel resistensi pada generasi F2 . Frekuensi alel resistensi diperkirakan menurut rumus yang diusulkan oleh Andow dan Alstad 36 :
![]()
di mana q merupakan estimasi frekuensi alel resistensi, S merupakan jumlah isoline positif, dan N merupakan jumlah isoline yang diuji.
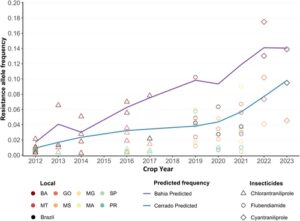
Frekuensi yang diestimasi oleh penyaringan F 2 digunakan untuk mengembangkan model prediktif berdasarkan data literatur yang tersedia untuk memantau frekuensi alel resistensi terhadap diamida di S. frugiperda di Brasil. 37 – 39 Dengan menggunakan basis data estimasi frekuensi alel untuk diamida dan populasi lapangan FAW dari tahun 2012 hingga 2022, dua model prediktif disorot untuk menunjukkan tren peningkatan frekuensi: satu untuk wilayah Bahia (skenario terburuk) dan satu lagi untuk semua wilayah di Cerrado Brasil. Untuk mengeksplorasi tren evolusi resistensi, kami menerapkan GLMM yang menggabungkan musim panen sebagai efek tetap dan variabilitas tahunan sebagai efek acak, dengan istilah penghalusan mengikuti struktur model GAM. 42 Semua analisis dan visualisasi dilakukan dengan menggunakan perangkat lunak R versi 4.3.1. 43
2.4 Pewarisan ketahanan Spodoptera frugiperda terhadap cyantraniliprole
Uji hayati dengan pola makan dilakukan setelah 10 generasi galur CYA-R di laboratorium untuk mengkarakterisasi dasar genetik resistensi. Pupa dari galur SUS dan CYA-R dipisahkan berdasarkan jenis kelamin dan disimpan dalam gelas plastik (50 mL). Saat dewasa muncul, persilangan timbal balik antara 25 pasangan (♀CYA-R × ♂SUS dan ♀SUS × ♂CYA-R) disimpan dalam kandang (tabung PVC berdiameter 10 cm × tinggi 20 cm). Keturunan heterozigot dari persilangan resiprokal (F 1 ) dikenakan uji hayati hamparan diet untuk melakukan kurva respons konsentrasi untuk siantraniliprole dan memperkirakan LC 50 untuk setiap galur heterozigot menggunakan konsentrasi logaritmik yang diberi jarak antara 1,80 dan 180 μg ai mL −1 siantraniliprole.
Kemungkinan resistensi yang terkait dengan jenis kelamin atau efek maternal diverifikasi dengan menganalisis hasil yang diperoleh dalam persilangan resiprokal. Tingkat dominasi resistensi rata-rata diperkirakan dengan membandingkan LC50 dari keturunan F1 dari galur heterozigot dengan LC50 dari galur parental menggunakan rumus yang diusulkan oleh Stone 44 :
![]()
di mana D adalah derajat dominasi rata-rata, dan X SS , X RR , dan X RS = log 10 (LC50 ) dari galur SUS, CYA-R, dan heterozigot H1 dan H2 , berturut-turut. Nilai D bervariasi antara -1 dan 1, dengan resistensi menjadi dominan sepenuhnya ketika D = 1, dominan tidak lengkap ketika 0 < D < 1, resesif tidak lengkap ketika -1 < D < 0, dan resesif sepenuhnya ketika D = -1. Sebagai pelengkap metode yang diusulkan oleh Stone, 44 data mortalitas dari bioassay diajukan ke rumus yang diusulkan oleh Bourguet et al . 45 untuk memverifikasi derajat dominasi pada berbagai konsentrasi:
![]()
di mana D adalah derajat dominasi rata-rata, dan M SS , M RR , dan M RS adalah mortalitas dari strain SUS, CYA-R, dan heterozigot. Nilai D yang mendekati nol menunjukkan pewarisan resesif sepenuhnya, sedangkan nilai yang mendekati satu menunjukkan pewarisan dominan sepenuhnya.
Pewarisan dianalisis untuk sifat monogeniknya menggunakan uji 𝜒 2 antara data mortalitas yang diamati dan yang diharapkan dari backcross. 46 Mortalitas yang diharapkan dari backcrossing pada konsentrasi tertentu dihitung berdasarkan mortalitas rata-rata antara keturunan F 1 dan orang tua mereka yang diusulkan oleh Sokal dan Rohlf 47
![]()
di mana Ni = Mortalitas yang diamati dari persilangan balik keturunan dengan konsentrasi i, p adalah proporsi mortalitas yang dihitung dengan model Mendelian (p = ( a + b ) / 2), ni adalah jumlah individu keturunan yang diuji pada konsentrasi i, q adalah proporsi mortalitas kelangsungan hidup yang diharapkan ( q = 1 – p ), a adalah mortalitas galur parental, dan b adalah mortalitas keturunan heterozigot F 1 . Hipotesis pewarisan monogenik ditolak ketika χ 2 yang dihitung ≥ χ 2 yang ditabulasikan dengan derajat kebebasan sama dengan 1 ( P < 0,05).
Untuk memperkirakan LC50 , digunakan model linear umum (GLM), dengan asumsi distribusi binomial dan fungsi hubungan probit. Kualitas kecocokan model dinilai menggunakan uji χ2 dan ‘ plot setengah normal’ dengan amplop simulasi dari paket hnp.48 Nilai LC50 diperkirakan menggunakan fungsi ‘dose.p’ dari paket MASS, 49 dan galur heterozigot dibandingkan menggunakan uji paralelisme dan kesetaraan dengan membandingkan model dengan interaksi lengkap dan model tereduksi menggunakan uji Chi-square. Semua analisis dan visualisasi dilakukan menggunakan perangkat lunak R versi 4.3.1.43
2.5 Resistensi silang antara cyantraniliprole dan insektisida diamida lainnya
Untuk memverifikasi resistensi silang S. frugiperda yang resistan terhadap cyantraniliprole dan flubendiamide (Belt®, SC 480 g ai l −1 , Bayer SA, São Paulo/SP, Brazil), chlorantraniliprole (Premio®, SC 200 g ai l −1 , FMC Quimica do Brasil Ltda., Campinas/SP, Brazil), dan cyclaniliprole (Goemon®, SL 50 g ai l −1 , ISK Biosciences do Brasil, Indaiatuba/SP, Brazil), kurva konsentrasi-respons diperoleh untuk setiap insektisida untuk strain SUS dan CYA-R. Metode bioassay sama seperti yang dijelaskan sebelumnya, dan mortalitas dinilai 96 jam setelah paparan. Ada atau tidaknya resistensi silang diverifikasi dengan membandingkan rasio resistensi (RR) dengan membagi LC50 strain CYA-R dengan LC50 strain SUS untuk setiap insektisida.
2.6 Analisis molekular dan genotipe reseptor ryanodine pada Spodoptera frugiperda yang resistan terhadap cyantraniliprole
Untuk menyelidiki dasar molekuler resistensi cyantraniliprole pada S. frugiperda , total RNA diekstraksi dari 30 larva instar ketiga awal dari strain SUS dan CYA-R menggunakan RNeasy® Plant Mini Kit (Qiagen). Sampel RNA diperlakukan dengan DNase I untuk menghilangkan DNA genom yang mengontaminasi (Thermo Scientific, Waltham, MA, AS), mengikuti protokol pabrik. Kuantitas dan integritas RNA dinilai menggunakan spektrofotometer NanoDrop (Thermo Scientific, Waltham, MA, AS) dan elektroforesis gel agarosa 1,0%. RNA dinormalisasi menjadi 1000 ng.μl −1 , dan 1 μg digunakan untuk mensintesis cDNA untai pertama menggunakan SuperScript® III First-Strand Synthesis System for RT-PCR (Invitrogen) dan primer Oligo(dT)20 (Promega, Madison, WI, AS).
Untuk mengurutkan reseptor ryanodine (RyR), primer yang menargetkan domain transmembran II hingga V dirancang (Tabel 2 ) berdasarkan wilayah C-terminal gen RyR.50 Primer mengamplifikasi dua fragmen 740 dan 1038 pasangan basa (bp). Amplifikasi PCR dilakukan dengan menggunakan DNA polimerase Q5 High-Fidelity (New England BioLabs). Setiap reaksi PCR 50 μL berisi 10 μL buffer, 1 μL dNTP, 0,5 μL DNA polimerase Q5, 2,5 μL setiap primer, dan 5 μL cDNA yang diencerkan 10x, dengan air MilliQ ditambahkan untuk mencapai volume akhir. Kondisi PCR adalah 35 siklus pada suhu 98 °C selama 10 detik, 60 °C selama 30 detik, dan 72 °C selama 30 detik, diikuti oleh ekstensi akhir pada suhu 72 °C selama 2 menit. Produk PCR dianalisis menggunakan elektroforesis gel agarosa 1%, dimurnikan menggunakan manik magnetik Beckman Coulter™ Agencourt AMPure XP (Beckman Coulter, Inc. Brea CA 92821, AS), dan selanjutnya dikirim untuk pengurutan Sanger. Hasil pengurutan Sanger diselaraskan dengan urutan referensi reseptor ryanodine P. xylostella (NP_001296002) untuk mengidentifikasi potensi mutasi.
| Bahasa Inggris: Primer | Urutan (5′ → 3′) |
|---|---|
| Bagian 1-F | ACGACGATGCACTAGAAG |
| Sf 2-F | GCCATCGAAGCTGAGAGCAA |
| Bagian 1-R | GTTCCTGTTGACCTCGTCGT |
Genotipe individual dilakukan pada 30 larva instar ketiga dari galur SUS dan CYA-R untuk memverifikasi keberadaan mutasi I4790K pada reseptor ryanodine. DNA genomik (gDNA) diekstraksi menggunakan Chelex 100 Resin (Bio-Rad, Hercules, CA, AS) pada 10%, dinormalisasi, dan digunakan dalam uji PCR kuantitatif waktu nyata (RT-qPCR). Reaksi dilakukan pada Sistem QuantStudio™ 5 (Thermo Scientific, Waltham, MA, AS) menggunakan kit TaqPath™ ProAmp Master Mix. Setiap reaksi qPCR 20 μL berisi 5 μL 3 ng.μl −1 gDNA, 10 μL TaqPath™ ProAmp Master Mix, 10 μM setiap primer, 0,4 μL probe spesifik mutasi, dan air MilliQ. Protokol qPCR melibatkan langkah denaturasi awal pada suhu 95 °C selama 5 menit, diikuti oleh 40 siklus pada suhu 95 °C selama 15 detik dan 60 °C selama 30 detik. Primer dan probe untuk mendeteksi mutasi I4790K dirancang seperti yang dijelaskan oleh Boaventura et al . 50 dan Okuma et al . 23 (Tabel 3 ).
| Urutan (5′ → 3′) | Orientasi | Pengubah | Referensi |
|---|---|---|---|
| CGACGATGCACTAGAAGTG | Maju | – | Mod. dari Boaventura dkk . 50 |
| ACCTTGAGATGGTAGTACC | Balik | – | Mod. dari Boaventura dkk . 50 |
| TGCTGCTAAACTCATCGGGT | Menguji | 5′ FAM + 3′ BHQ1 | Sf . mod I4790K. dari Okuma dkk . 23 |
| TGTCGCTCGCTATACTCATCG | Menguji | 5′ HEKSAGON + 3′ BHQ1 | Sf . I4790I mod. dari Boaventura et al . 50 |
3 HASIL
3.1 Pemantauan kerentanan Spodoptera frugiperda terhadap cyantraniliprole
Tingkat kelangsungan hidup rata-rata populasi lapangan S. frugiperda tetap di bawah 10% sejak pemantauan laboratorium dimulai pada tahun 2017. Namun, outlier diamati, dengan tingkat kelangsungan hidup antara 12,39% dan 14,03% di wilayah tertentu Goiás (GO) dan Bahia (BA) (Gbr. 1 (A) ). Tingkat kelangsungan hidup rata-rata pada musim panen 2022 secara signifikan lebih tinggi (3,88% ± 4,35) dibandingkan tahun sebelumnya (2,03% ± 4,41), yang menunjukkan variasi yang cukup besar di berbagai lokasi. Model campuran linier umum (GLMM) mengungkapkan bahwa tingkat kelangsungan hidup di BA lebih tinggi, diikuti oleh Mato Grosso (MT) dan GO (Gbr. 1(B) ) (Intercept = 0,0391 dan β = 0,0514; P = 0,0348).
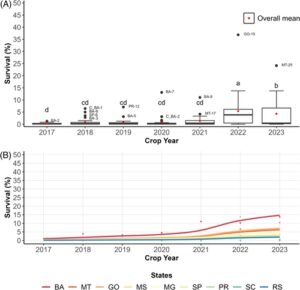
3.2 Layar F 2 untuk memperkirakan frekuensi alel resistensi
Frekuensi alel resistensi yang diperkirakan untuk cyantraniliprole ditentukan dalam tujuh populasi lapangan S. frugiperda , dengan 75.644 larva yang dinilai pada konsentrasi diskriminatif. Frekuensi alel resistensi yang diperkirakan tertinggi diamati di BA barat, di mana ia tetap konstan sepanjang musim panen 2022–2023, dengan frekuensi rata-rata 0,1442. Hal ini menunjukkan persistensi individu yang resisten di wilayah tersebut (Tabel 4 ). Di negara bagian GO dan MT, frekuensi rata-rata yang diperkirakan masing-masing adalah 0,0756 dan 0,0734, yang mewakili proporsi individu yang resisten (Wrr) hampir empat kali lebih rendah daripada di BA. Skenario paling tidak kritis diamati di Mato Grosso do Sul (MS), di mana frekuensi yang diperkirakan adalah 0,0408 (Tabel 4 ).
| Kode | Isoline awal | Isoline yang diuji | Serangga yang diuji | Isoline positif | Frekuensi (q) (CI 95%) |
|---|---|---|---|---|---|
| PERGI-19 | 190 | 123 | 14.107 | 50 | 0,1020 (0,0770–0,1299) |
| PERGI-22 | 189 | 103 | 12 816 | 18 | 0,0453 (0,0275–0,0671) |
| Goias (GO) | 379 | 226 | 26 923 | 68 | 0,0756 (0,0594–0,0936) |
| BA-9 | 186 | 69 | 8033 | 36 | 0,1303 (0,0937–0,1717) |
| Bahasa Indonesia: C_BA-6 | 154 | 48 | 5760 | 34 | 0,1750 (0,1257–0,2305) |
| BA-10 | tahun 201 | 115 | 13 568 | 64 | 0,1389 (0,1091–0,1716) |
| Bahia (BA) | 541 | 232 | 27 361 | 134 | 0,1442 (0,1224–0,1674) |
| MT-20 | 176 | 90 | 10 800 | 26 | 0,0734 (0,0490–0,1021) |
| Bahasa Indonesia: MS-14 | 184 | 90 | 10 560 | 14 | 0,0408 (0,0230–0,0632) |
| Cerrado (semua) | tahun 1280 | 638 | 75 644 | 243 | 0,0949 (0,0838–0,1065) |
3.3 Pewarisan ketahanan Spodoptera frugiperda terhadap cyantraniliprole
Galur resistan yang dipilih dengan metode penyaringan F 2 (CYA-R) menunjukkan LC 50 yang diperkirakan sebesar 10095,31 μg ai siantraniliprole mL −1 , yang menunjukkan rasio resistensi sebesar 3414 kali lipat (Tabel 5 ). Kedua heterozigot menunjukkan sedikit variasi dalam nilai LC 50 yang diperkirakan , menghasilkan rasio resistensi sebesar 3,69 kali lipat untuk H 1 (♂CYA-R × ♀SUS) dan 3,02 kali lipat untuk H 2 (♂SUS × ♀CYA-R) (Tabel 5 ). Temuan-temuan ini menunjukkan bahwa pewarisan kemungkinan bersifat autosomal, karena heterozigot menunjukkan rasio resistensi yang secara kualitatif serupa antara kedua galur berdasarkan uji paralelisme (χ 2 = 1,22, df = 1, P = 0,27), meskipun tidak sama (χ 2 = 6,43, df = 2, P = 0,04). Selain itu, heterozigot menunjukkan tingkat resistensi yang lebih dekat dengan galur SUS, seperti yang tercermin dari rasio resistensi yang rendah, yang menunjukkan pola pewarisan resesif (Gbr. 2 ). Hasil ini selanjutnya didukung oleh nilai negatif derajat dominasi (D) yang dihitung menggunakan rumus Stone 44 (Tabel 5 ) (Gbr. 3 ). Pada konsentrasi diagnostik (180 μg ai siantraniliprole mL −1 ), hasil dominasi menunjukkan bahwa pola pewarisan sepenuhnya resesif (Gbr. 3 ).
| Ketegangan | N † | LC 50 ‡ (μg ai mL −1 ) (CI 95%) | χ 2 (df) § | P | Kemiringan (± Tenggara) | RR 50 (CI 95%) ‖ | D ¶ |
|---|---|---|---|---|---|---|---|
| SUS | 804 | 2,96 (2,56–3,42) | 10.24 (6) | 0.12 | 2,01 (± 0,14) | – | – |
| H 1 (♂CYA-R × ♀ SUS) | tahun 907 | 10,92 (9,67–12,33) | 9.60 (7) | 0.21 | 2,26 (± 0,14) | 3,69 (3,07–4,44) | -0,68 |
| H2 ( ♂SUS ×♀CYA-R) | 941 | 8.93 (8.00–9.96) | 6.30 (7) | 0.51 | 2,51 (± 0,17) | 3.02 (2,49–3,66) | -0,73 |
| CYA-R (F 11 ) | 683 | 10 095.31 (9216.24–11 058.23) | 8.33 (6) | 0.21 | 2,99 (± 0,20) | 3414.29 (2808–4141) | – |
† Jumlah serangga yang diuji.
‡ Konsentrasi mematikan dari larutan insektisida yang digunakan yang membunuh 50% individu, dengan interval kepercayaan 95%.
§ Uji chi-square (derajat kebebasan).
‖ Rasio resistensi (RR) diperoleh dengan membagi LC50heterozigot (H1/H2) dan resisten (CYA-R) dengan LC50strain rentan (SUS).
¶ Tingkat dominasi diperoleh dengan menggunakan rumus Stone (1968). 44
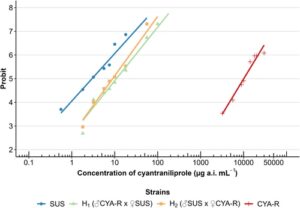
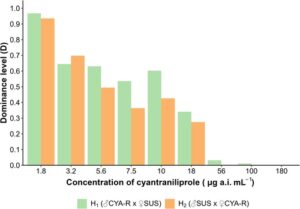
Hipotesis pewarisan monogenik diterima dengan membandingkan hasil mortalitas yang diamati dengan data mortalitas yang diharapkan dari model Mendel menggunakan uji chi-square (Tabel 6 ).
| Konsentrasi | ♂ F 1 × ♀ CYA-R | ♂ CYA-R × ♀ F 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| (μg.a.i.mL −1 ) | Catatan † | Eksp ‡ | χ 2 § | P ‖ | Catatan | Eksp | χ 2 | P |
| pukul 18.00 | 40 | 33 | 0.82 | 0.37 | 33 | 33 | 0,05 | 0.82 |
| 56.00 | 41 | 49 | 2.50 | 0.11 | 51 | 49 | 0.24 | 0.63 |
| Rp 100.000 | 52 | 50 | 0.21 | 0,65 | 50 | 50 | 0.00 | 1.00 |
| Rp. 180.000 | 53 | 50 | 0.42 | 0.52 | 53 | 50 | 0.53 | 0.47 |
† Angka kematian yang diamati (%). ‡ Angka kematian yang diharapkan (%) menurut pewarisan monogenik Mendel. § Hasil chi-kuadrat. ‖ Tingkat signifikansi probabilitas 5% (χ2 >3,84, df = 1,P < 0,05).
3.4 Resistensi silang antara cyantraniliprole dan insektisida diamida lainnya
Resistensi silang dikonfirmasi antara siantraniliprole dan semua diamida lain yang diuji. Hasil kami menunjukkan bahwa untuk dua diamida pertama yang dikomersialkan, flubendiamide dan klorantraniliprole, rasio resistensi masing-masing >229.664 kali lipat dan 112.359 kali lipat, yang menunjukkan bahwa bahkan formulasi komersial yang tidak diencerkan tidak cukup untuk mencapai angka kematian 50%. Rasio resistensi untuk siklaniliprole juga signifikan, yaitu 19.283 kali lipat, yang menunjukkan bahwa galur yang resistan terhadap siantraniliprole menunjukkan resistensi silang yang tinggi terhadap semua diamida yang diuji di Brasil (Tabel 7 ).
| Insektisida | Ketegangan | N † | LC 50 ‡ (μg ai mL −1 ) (CI 95%) | χ 2 (df) § | P | Kemiringan (± Tenggara) | RR 50 ‖ |
|---|---|---|---|---|---|---|---|
| Flubendiamida | SUS | 600 | 2,09 (1,67–2,61) | 6.21 (5) | 0.28 | 1,25 (± 0,11) | – |
| CYA-R | 624 | >480.000,00 | – | – | – | >229 664.00 | |
| Klorantraniliprol | SUS | 792 | 1,78 (1,51–2,11) | 6.13 (6) | 0.41 | 1,67 (± 0,11) | – |
| CYA-R | 827 | >200.000,00 | – | – | – | >112 359.00 | |
| Siklaniliprol | SUS | 669 | 0,12 (0,10–0,15) | 1.38 (6) | 0,96 | 1,52 (± 0,10) | – |
| CYA-R | 876 | 2488.55 (2199.16 – 2816.01) | 11.67 (9) | 0.23 | 2.10 (± 0,12) | 19 283.67 |
† Jumlah serangga yang diuji. ‡ Konsentrasi mematikan dari larutan insektisida yang digunakan yang membunuh 50% individu, dengan interval kepercayaan 95%. § Uji chi-square (derajat kebebasan). ‖ Rasio resistensi (RR) diperoleh dengan membagi LC50strainresisten (CYA-R) dengan LC50strainrentan (SUS).
3.5 Prediksi evolusi resistensi insektisida diamides terhadap S. frugiperda
Ketika membandingkan siantraniliprole dengan diamida lain, seperti flubendiamide dan klorantraniliprole, yang dirilis pada pertengahan tahun 2010, tingkat kelangsungan hidup untuk diamida yang lebih tua ini tetap di bawah 10% selama 4 tahun pertama pemantauan (2014–2018), kecuali untuk BA (Gbr. 4 ). Di wilayah dengan sistem produksi pertanian intensif, termasuk Negara Bagian Cerrado Brasil (BA, MT, GO, MS, dan MG), ada sedikit peningkatan dalam tingkat kelangsungan hidup. Populasi lapangan dari Brasil selatan (Paraná-PR, Santa Catarina-SC, dan Rio Grande do Sul-RS) secara konsisten menunjukkan tingkat kelangsungan hidup rata-rata di bawah 10% untuk semua diamida, yang menunjukkan kerentanan yang stabil.
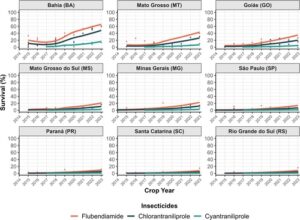
Tren peningkatan keseluruhan dalam frekuensi alel resistensi yang diestimasikan diamati di seluruh musim tanam di Brasil (Gbr. 5 ). Model prediktif yang disesuaikan menggunakan GLMM menggabungkan data kami tentang cyantraniliprole bersama dengan informasi dari Ribeiro, 38 Bolzan, 37 dan Okuma et al ., 39 mengenai chlorantraniliprole dan flubendiamide yang menunjukkan peningkatan yang konsisten dalam alel resistensi untuk diamides dari waktu ke waktu. BA menyoroti peningkatan yang signifikan dalam frekuensi alel resistensi dibandingkan dengan rata-rata Cerrado, yang menunjukkan tekanan selektif yang lebih kuat di wilayah ini. Peningkatan dalam evolusi resistensi di BA mungkin disebabkan oleh praktik pertanian lokal (tiga musim tanam dan lahan subur yang luas) yang mengintensifkan tekanan ini dan meningkatkan penyebaran alel resistan.
3.6 Analisis molekular dan genotipe reseptor ryanodine pada Spodoptera frugiperda yang resistan terhadap cyantraniliprole
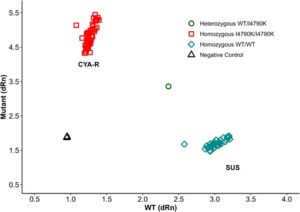
Pengurutan domain transmembran reseptor ryanodine pada galur CYA-R mengungkap substitusi timin (T) oleh adenosin (A), yang menghasilkan lisin (K) asam amino yang menggantikan isoleusin (I) pada posisi 4734 di S. frugiperda , yang sesuai dengan mutasi I4790K di P. xylostella (NP_001296002) (Gbr. 6 ). Genotipe 30 larva yang diisolasi dari galur yang resistan terhadap siantraniliprole (CYA-R) menunjukkan bahwa 100% individu bersifat homozigot untuk mutasi I4790K (Gbr. 7 ). Karena sebagian besar mutasi terkait resistensi pada reseptor ryanodine telah dijelaskan di wilayah transmembran ini, hasil kami menunjukkan bahwa mutasi I4790K berperan dalam resistensi siantraniliprole. Namun, mekanisme potensial lainnya tidak dapat dikesampingkan karena belum dieksplorasi dalam penelitian ini.



Leave a Reply