
Ringkasan
- Perubahan iklim akibat manusia diperkirakan akan membawa kekeringan yang lebih sering dan suhu yang lebih tinggi di Amerika Serikat bagian barat, yang mengancam hutan aspen gemetar yang penting secara ekologis.
- Kami menggunakan kurva kerentanan spesifik cincin cabang aspen di sepanjang dua gradien iklim untuk menentukan apakah kerusakan pada membran lubang terakumulasi seiring bertambahnya usia xilem.
- Kami menemukan bahwa cincin yang lebih tua dari 3 tahun memiliki penurunan yang signifikan dalam konduktivitas hidrolik, terutama pada potensi air musim panas rata-rata untuk spesies tersebut. Perbedaan ini bukan disebabkan oleh perbedaan diameter pembuluh, tetapi perbedaan dalam seberapa banyak xilem yang aktif antara cincin yang lebih tua dari 3 tahun dan 1 tahun, yang menunjukkan adanya kerusakan yang terakumulasi pada membran lubang yang mengganggu transportasi air.
- Kerentanan terhadap emboli berbeda-beda di setiap umur lingkaran dan antara populasi yang lebih basah dan lebih kering, menggarisbawahi bahwa kerusakan akibat kekeringan dapat terakumulasi hingga ke tingkat yang mematikan jika xilem tidak beraklimatisasi terhadap perubahan iklim pada pertumbuhan yang lebih baru.
Perkenalan
Hutan memainkan peran penting dalam sistem Bumi, karena mereka berdampak pada siklus karbon, air, dan energi serta iklim (Bonan, 2008 ). Namun, hutan telah terancam karena perubahan iklim antropogenik, yang telah membawa peristiwa kekeringan yang lebih sering bersamaan dengan meningkatnya permintaan kelembaban atmosfer karena suhu yang lebih tinggi (Williams et al ., 2013 ; IPCC, 2014 ; Spinoni et al ., 2018 ; Grossiord et al ., 2020 ). Kekeringan jenis perubahan iklim ini telah mengakibatkan kerusakan pada xilem yang membatasi pertumbuhan pohon selama bertahun-tahun setelah kekeringan atau menyebabkan peristiwa kematian (Breshears et al ., 2005 ; Anderegg et al ., 2013b , 2015 ), yang mengancam peran hutan sebagai penyerap karbon (Allen et al ., 2010 ). Perubahan yang disebabkan oleh perubahan iklim dalam rezim kekeringan menyoroti perlunya memahami dampak jangka panjang dari stres kekeringan pada transportasi hidrolik tanaman. Khususnya, apakah kerusakan terakumulasi hingga tingkat yang mematikan atau apakah xilem berpotensi pulih dan bahkan bergeser melalui pertumbuhan baru untuk menjadi terbiasa dan lebih tahan terhadap kekeringan adalah hal yang tidak diketahui secara pasti (Corcuera et al ., 2004 ; Jacobsen & Pratt, 2014 ; Tomasella et al ., 2019 ; Anderegg et al ., 2020 ; Pratt et al ., 2020 ; Pritzkow et al ., 2021 ).
Fisiologi respons xilem terhadap beberapa kekeringan itu rumit dan kurang dipahami. Peristiwa kekeringan parah yang menyebabkan paparan terus-menerus terhadap stres kekeringan dapat menyebabkan kerusakan xilem yang meningkatkan kerentanannya terhadap kekeringan, (yaitu kelelahan kavitasi) pada beberapa spesies (Hacke et al ., 2001 ; Feng et al ., 2021 ). Selama stres kekeringan, tegangan xilem dapat melebihi toleransi saluran karena berdampak pada membran di dalam lubang saluran. Ketika ini terjadi, gas ditarik ke dalam saluran dan menciptakan emboli, yang menghalangi kemampuan saluran untuk mengangkut air. Emboli dapat menyebar dari satu saluran ke saluran lain melalui membran lubang. Kelelahan kavitasi terjadi ketika kerusakan permanen pada membran lubang terjadi dan menjadi kurang tahan terhadap penyemaian udara (yaitu lubang ‘bocor’), bahkan jika saluran diisi ulang (Hacke et al ., 2001 ). Lebih jauh lagi, penuaan xilem juga dapat memengaruhi resistensi emboli dan konduktansi air dalam jangka waktu yang panjang (Sperry et al ., 1991 ). Hebatnya, sejauh pengetahuan kami, belum banyak penelitian tentang penuaan xilem dan perubahan kerentanan dalam cincin individu dari waktu ke waktu, karena hampir semua penelitian memeriksa respons batang penuh atau dalam bagian besar xilem batang (Domec & Gartner, 2001 ; Spicer & Gartner, 2001 ; Delzon et al ., 2004 ; Ford et al ., 2004 ; tetapi lihat Melcher et al ., 2003 , untuk contoh penelitian yang menggunakan pola kerentanan spesifik pembuluh, Domec & Gartner, 2002 , untuk pola antara kayu awal dan kayu akhir, dan Venturas et al ., 2019 , untuk perbandingan antara xilem primer dan sekunder), dan dengan demikian, variasi dalam kerentanan dan konduktivitas di seluruh cincin spesifik tidak dipahami dengan baik. Perbandingan awal pada batang dengan umur berbeda memang menghasilkan konduktivitas yang bervariasi, tetapi pola ini ditemukan pada spesies berpori cincin dan mungkin bervariasi pada tipe anatomi kayu yang berbeda (Pratt et al ., 2020 ).
Amerika Serikat bagian barat telah mengalami beberapa kekeringan parah akibat ‘perubahan iklim’ dalam beberapa dekade terakhir, yang menjadikannya tempat yang ideal untuk memeriksa respons hidrolika tanaman terhadap beberapa kekeringan (Breshears et al ., 2005 ; Williams et al ., 2013 ; Zhang et al ., 2021 ). Kekeringan parah ini telah memengaruhi beberapa spesies pohon dominan, termasuk hutan aspen gemetar ( Populus tremuloides Michx.) yang penting secara ekologis. Dalam 15 tahun terakhir, hutan aspen telah mengalami peristiwa kematian yang meluas yang dikenal sebagai penurunan aspen mendadak (SAD; Worrall et al ., 2008 ; Anderegg et al ., 2013a , b ). SAD tampaknya terkait dengan kerusakan yang terakumulasi selama beberapa tahun pada xilem karena kelelahan kavitasi (Anderegg et al ., 2013b ; Feng et al ., 2021 ). Hutan yang telah mengalami SAD mungkin lebih berisiko di masa mendatang terhadap kekeringan yang tidak terlalu parah (Anderegg et al ., 2013b ), pola yang sering terlihat pada angiospermae berkayu lain yang terpapar stres kekeringan (DeSoto et al ., 2020 ). Selain itu, emboli beku-cair juga dapat berinteraksi dengan stres kekeringan untuk meningkatkan kerentanan xilem (Sperry et al ., 1994 ; Langan et al ., 1997 ; Feng et al ., 2015 ). Peristiwa pembekuan dapat meningkatkan ketegangan latar belakang yang diberikan pada sistem pengangkutan air, yang menyebabkan ‘kelelahan beku’ tambahan (Feng et al ., 2015 ). Ukuran saluran memengaruhi kerentanan emboli beku-cair dan emboli kekeringan dan oleh karena itu dapat berguna dalam menunjukkan perbedaan dalam konduktivitas dan kerentanan terhadap kedua jenis stresor xilem (Langan et al ., 1997 ).
Kami menguji apakah xilem menunjukkan akumulasi kerusakan dari beberapa kekeringan parah di hutan aspen di Amerika Serikat bagian barat. Kami bertujuan untuk menjawab tiga pertanyaan utama menggunakan data hidrolik dan anatomi sepanjang beberapa gradien iklim: (1) Bagaimana cincin usia yang berbeda berbeda dalam kemampuannya untuk menahan emboli? Kami berhipotesis bahwa cincin yang lebih tua akan lebih rentan terhadap emboli; (2) bagaimana cincin usia yang berbeda berbeda dalam konduktivitas hidrolik dan luas area xilem aktif? Kami berhipotesis bahwa cincin yang lebih baru akan memiliki lebih banyak konduktivitas dan persentase area konduktif aktif yang lebih tinggi; (3) bagaimana populasi aspen dari rezim kekeringan yang berbeda berbeda dalam akumulasi kerusakannya? Kami berhipotesis bahwa aspen dari daerah dengan rezim kekeringan yang lebih parah akan mengalami peningkatan kelelahan kavitasi dan dengan demikian meningkatkan kerentanan terhadap emboli.
Bahan dan Metode
Ringkasan
Kami menggunakan kurva kerentanan hidrolik untuk mengukur kerentanan xilem terhadap emboli di Populus tremuloides (Michx.). Sebagian besar kurva kerentanan dibuat menggunakan seluruh luas penampang sampel; namun, kurva ini tidak mempertimbangkan perbedaan di antara cincin pertumbuhan tahunan dan mungkin melebih-lebihkan kontribusi cincin tua terhadap konduktansi batang secara keseluruhan (Melcher et al ., 2003 ; Fukuda et al ., 2015 ). Kemungkinan besar penurunan tajam di awal kurva kerentanan disebabkan oleh emboli pada cincin yang lebih tua (Fukuda et al ., 2015 ). Untuk menguji pertanyaan yang dijelaskan sebelumnya, kami melakukan kurva kerentanan khusus cincin dan kami memasangkannya dengan pengukuran anatomi dan perfusi pewarna untuk menentukan daerah fungsional xilem dalam populasi aspen di dua gradien iklim.
Sistem belajar
Kami menggunakan dua gradien iklim komplementer: gradien iklim elevasi (dalam populasi) dan gradien iklim lintas populasi – untuk mengeksplorasi pertanyaan studi utama kami. Gradien elevasi berada di Colorado Barat Daya di dalam Hutan Nasional San Juan (SJNF). Kami menggunakan tegakan hutan yang menghadap ke selatan yang terletak di tiga pita elevasi (rendah, sedang, dan tinggi) yang mencakup rentang elevasi dari 2600 m hingga 3200 m, dipisahkan setiap 300 m (Anderegg & Hillerislambers, 2016 ). Sepanjang gradien ini, presipitasi meningkat dan suhu menurun seiring dengan elevasi (Tabel Informasi Pendukung S1 , S2 ). Kami mengumpulkan sampel dari gradien elevasi San Juan dari tiga pita elevasi pada 8 Juli dan 23 Agustus 2021. Kedekatan tegakan aspen di SJNF memungkinkan kami untuk membandingkan kemungkinan dalam populasi yang sama dengan tingkat aliran gen yang tinggi.
Kami menggunakan gradien iklim multipopulasi untuk membandingkan berbagai populasi genetik (Kerr et al ., 2023 ). Kami mengumpulkan sampel dari lima populasi aspen yang mencakup gradien iklim di Hutan Nasional Dixie, San Juan, Uncompahgre, White River, dan Uinta di Colorado dan Utah, AS, dalam bulan Juni hingga Agustus 2022 (Tabel S3 ). Kami memberi peringkat lokasi dalam urutan yang tercantum di atas karena lokasi tersebut mewakili gradien dari paling selatan ke paling utara dan bergradasi dari panas dan kering ke hangat dan basah (Tabel S3 ). Semua plot memiliki kemiringan, aspek (Selatan), dan elevasi yang sama ( c . 3000 m) seperti yang dijelaskan dalam Kerr et al . ( 2023 ), kecuali untuk lokasi White River, yang tidak dapat diakses karena kondisi jalan. Sebagai gantinya, kami membuat plot White River baru dengan elevasi yang sedikit lebih rendah ( c . 2665 m) dengan aspek Timur Laut. Kombinasi kedua gradien ini memberikan pengujian pelengkap atas pertanyaan kami dan memungkinkan kami untuk melihat peran potensial perbedaan tingkat populasi (yaitu genetik) dalam pola aklimatisasi dan akumulasi. Nilai indeks fiksasi ( F st ) untuk gradien iklim berkisar antara 0,04 hingga 0,12 sebagaimana ditentukan oleh Kerr et al . ( 2023 ), sementara satu populasi genetik dari gradien elevasi diasumsikan ( F st = 0).
Pengambilan sampel dan pengukuran
Di semua lokasi, kami mengumpulkan cabang bercabang dari tajuk tengah yang terpapar sinar matahari menggunakan senapan laras ganda kaliber 20 atau pemangkas tiang ( n = 9 untuk setiap lokasi di sepanjang gradien elevasi dan n = 6 untuk setiap lokasi di sepanjang gradien iklim). Kami segera membungkus ujung yang dipotong dengan tisu basah, menyegel cabang dalam kantong plastik, dan meletakkan cabang yang dibungkus di atas es untuk disimpan selama perjalanan kembali ke laboratorium. Keesokan harinya di laboratorium, kami memisahkan kedua sisi cabang bercabang dan membuang setidaknya 5 cm jaringan dari ujung potongan awal menggunakan silet.
Satu sisi dari setiap cabang bercabang digunakan untuk pengukuran hidraulik. Dengan menggunakan pisau cukur di bawah air, kami secara bertahap memotong cabang berdiameter 5 hingga 10 mm hingga panjangnya 14 cm. Kami secara langsung mengukur konduktivitas hidraulik asli ( K native ) menggunakan peralatan hidraulik seperti yang dijelaskan dalam Sperry et al . ( 1988a ), dengan larutan KCl 10 mM yang telah didegaskan, 0,2 μm. Konduktivitas ( K h ) diukur untuk seluruh luas penampang (semua), cincin yang lebih tua dari 3 tahun (> 3), cincin berusia 2 hingga 3 tahun (2–3), dan cincin terbaru (baru; Gambar 1 ) menggunakan Persamaan 1 :
![]()
dimana Δ
adalah perbedaan tekanan antara pasokan air ke batang dan tinggi permukaan air pada skala. Selain itu, suhu larutan KCl dikoreksi ke resistansi 20°C (Yang & Tyree, 1993 ). Kami menggunakan tusuk gigi untuk mengoleskan lem sianoakrilat (lem Serba Guna Krazy Glue; Newell Office Brands, Atlanta, Georgia, AS) untuk memblokir cincin tertentu dan menghitung konduktivitas spesifik area xilem ( K s ) dengan membagi K h dengan area untuk setiap cincin (Gbr. S1 ). Penggunaan lem sianoakrilat telah terbukti sebagai metode yang efektif untuk memblokir konduktansi air dalam penelitian sebelumnya (Rodriguez-Zacarro et al ., 2019 ). Lem dioleskan ke satu ujung yang dipotong dalam beberapa detik dan kemudian dicelupkan ke dalam air sebelum merekatkan sisi lainnya. Setelah mengumpulkan pengukuran asli, kami memotong lem super di kedua ujungnya dan merendam sampel dalam 10 mmol larutan KCl dalam ruang vakum semalaman untuk menghilangkan emboli yang ada. Setelah infiltrasi vakum, kami mengukur ulang konduktivitas hidrolik maksimum ( K maks ) pada setiap wadah cincin menggunakan teknik perekatan yang sama seperti yang dijelaskan di atas. Kami melakukan pengukuran khusus cincin ini pada batang yang sama menggunakan teknik lem ini untuk mengurangi variasi di berbagai batang dan menghindari bias yang mungkin timbul dalam analisis karena penggunaan batang yang berbeda (Hacke et al ., 2022 ).
Kurva kerentanan dapat digunakan untuk mengukur resistensi emboli xilem secara efisien, seperti yang dijelaskan dalam Tobin et al . ( 2013 ) dan Venturas et al . ( 2016 ). Untuk membuat kurva kerentanan khusus cincin, sampel kami diputar dalam sentrifus pada kecepatan yang berkorelasi dengan tekanan negatif berikut: -0,5, -1, -2, -4, -8, -10 MPa, atau hingga batang mencapai setidaknya 90% pengurangan konduktivitas dari maksimum. Sebelum diputar dalam sentrifus, kami memotong lem di kedua ujungnya dengan silet. Setelah sentrifugasi, kami merekatkan kembali wadah cincin dan mengukur konduktivitas hidrolik untuk setiap wadah cincin. Kami menghitung persentase kehilangan konduktivitas (PLC) untuk setiap bin di setiap sampel relatif terhadap konduktivitas pada −0,5 MPa (koreksi kelelahan; Hacke et al ., 2001 ; Persamaan 2 ).
![]()
Kami kemudian menggunakan data PLC untuk membuat kurva kerentanan untuk setiap bin cincin dan setiap populasi dan lokasi dalam gradien elevasi, dengan menghilangkan kurva dengan nilai PLC negatif dari analisis. Tekanan pada kehilangan konduktivitas sebesar 50% ( P 50 ) dihitung dengan cara menyesuaikan setiap kurva kerentanan dengan kurva kecocokan Weibull dua parameter (Hacke et al ., 2015 ).
Bahasa Indonesia: Setelah menyelesaikan pengukuran kerentanan xilem, kami menggunakan pisau cukur berlapis Teflon (pisau GEM single-edge stainless steel berlapis Teflon; Electron Microscopy Sciences, Haffield, PA, AS) untuk membuat penampang cabang tipis di bagian bawah 2 cm sampel hidrolik. Kami mencitrakan penampang dengan mikroskop bedah (AmScope Outlet 7X-45X Trinocular Stereo Zoom Microscope, dengan Kamera Digital Mikroskop terpasang MU500; AmScope, Irvine, CA, AS), dan mengukur luas cincin dengan perangkat lunak analisis ImageJ (Schneider et al ., 2012 ) atau dengan perangkat lunak analisis AmScope (v.3.7.7934; AmScope). Kami membagi konduktivitas dengan luas cincin ini untuk menghitung konduktivitas hidrolik spesifik luas xilem ( K s ).
Sisi lain dari cabang bercabang yang dikumpulkan digunakan untuk perfusi pewarna untuk menentukan persentase area xilem aktif. Kami memangkas bagian berdiameter 5 hingga 10 mm di bawah air hingga setidaknya 10 cm panjangnya menggunakan silet. Mengikuti protokol pewarnaan pewarna aktif yang diuraikan dalam Jacobsen et al . ( 2007 ), kami menarik pewarna Safranin O 2,0 pH 0,5% berat/volume di bawah tekanan c . −2 kPa ke dalam segmen cabang selama 10 menit atau lebih lama hingga pewarna terlihat keluar melalui ujung potongan distal, yang kemudian segera dikantongi dan ditempatkan dalam freezer hingga pemotongan lebih lanjut. Kami menggunakan silet berlapis Teflon untuk membuat bagian tegak lurus ke cabang pada jarak 5 cm dari ujung potongan proksimal dan segera mengambil gambar bagian di bawah mikroskop bedah. Dengan menggunakan ImageJ , kami mengukur luas xilem yang berkaitan dengan setiap wadah, dan juga luas total pewarna untuk setiap wadah, yang memungkinkan kami menghitung persentase luas xilem aktif untuk setiap wadah cincin.
Bahasa Indonesia: Untuk menentukan apakah perbedaan konduktivitas di antara cincin dan populasi didorong oleh ukuran diameter pembuluh, kami mengukur diameter pembuluh dan diameter hidrolik dari setiap tempat sampah dan setiap populasi dalam gradien iklim. Ini diukur pada sampel yang sama dengan pengukuran konduktivitas hidrolik yang dilakukan untuk gradien iklim. Kami menggunakan silet berlapis Teflon untuk membuat penampang tipis yang kami pasang pada slide kaca dengan gliserol. Kami mengambil gambar dengan kamera (7.2 Color Mosaic Camera; SPOT Imaging, Sterling Heights, MI, AS) yang dipasang pada mikroskop (Nikon Eclipse E600W, Model RT KE; Diagnostic Instruments, Salt Lake City, UT, AS) pada perbesaran 200x, dan mengukur luas sedikitnya 100 pembuluh untuk setiap tempat sampah menggunakan perangkat lunak ImageJ . Kami mengubah luas pembuluh menjadi diameter pembuluh (dengan asumsi bahwa pembuluh itu berbentuk lingkaran). Kami juga menghitung diameter hidrolik menggunakan Persamaan 3 .
![]()
Kami mengumpulkan cabang-cabang tambahan dari pohon yang sama di populasi Uinta ( n = 5) dan White River ( n = 6) pada musim panas 2022 untuk menguji apakah pergerakan radial air melintasi cincin hadir. Ini dilakukan untuk menentukan apakah pemblokiran transportasi air dengan lem akan berhasil atau apakah air hanya akan bergerak melintasi batas cincin dan di sekitar penyumbatan lem. Kami menerapkan lem sianoakrilat ke ujung proksimal setiap bagian yang menutupi semua kecuali tiga cincin terbaru, kemudian melakukan perfusi pewarna seperti yang dijelaskan di atas. Kami menggunakan pisau cukur berlapis Teflon untuk memotong sampel beku, menjaga bagian dengan lem yang dioleskan, bagian langsung di bawah lem yang dioleskan, dan bagian 5 cm dari ujung proksimal. Kami berhati-hati untuk menjaga bagian yang dipotong dalam orientasi yang sama agar mudah menyelaraskan xilem. Bagian-bagian ini ditempatkan pada slide kaca dan dipasang dengan gliserol, segera dibayangkan dengan mikroskop bedah, dan dianalisis menggunakan perangkat lunak ImageJ . Untuk analisis gambar, kami mengidentifikasi irisan xilem yang bening, mudah untuk membedakan berapa banyak cincin yang diwarnai, dan mudah untuk disejajarkan untuk semua bagian. Untuk bagian 5 cm di atas ujung proksimal, kami mengukur total luas xilem dari cincin yang tidak direkatkan, selain luas pewarna yang meluas melewati daerah yang direkatkan. ‘Kebocoran’ pewarna (yaitu pergerakan radial pewarna ke dalam cincin yang direkatkan) dihitung sebagai persentase dari total luas yang direkatkan.
Bahasa Indonesia: Untuk memahami apakah penyumbatan xilem utamanya adalah emboli atau penyumbatan permanen, kami melakukan uji pewarna pada cabang asli yang dipintal dengan vakum dan berpasangan. Kami mengumpulkan cabang untuk uji pewarna ini pada 23 Agustus 2021, dari pita elevasi terendah dan tertinggi di SJNF ( n = 3 untuk setiap pita elevasi). Dua cabang dikumpulkan dari setiap pohon, dan dua segmen dari setiap cabang dipotong dan dipasangkan. Untuk satu cabang, kami mewarnai satu bagian dengan Pewarna Safranin O dalam kondisi asli, sementara bagian cabang lainnya diinfiltrasi vakum dengan 10 mmol larutan KCl semalaman, lalu diwarnai. Untuk cabang kedua, kami menginfiltrasi vakum kedua bagian semalaman, dan mewarnai satu bagian setelah pembilasan, sementara yang lain dipintal dalam sentrifus pada −2 MPa selama 6 menit sebelum pewarnaan. Setiap bagian pewarna setidaknya sepanjang 6 cm, dengan sampel yang disentrifugasi setidaknya sepanjang 10 cm agar pas di rotor (dengan spacer khusus untuk rotor sepanjang 14 cm). Kami memotong batang dengan pisau cukur berlapis Teflon pada jarak 5 cm dari ujung proksimal setiap bagian dan mengukur area xilem yang diwarnai dan total dengan perangkat lunak analisis Amscope , yang kemudian kami gunakan untuk menghitung persentase xilem yang diwarnai (aktif) untuk setiap bagian. Jika tidak ada perbedaan dalam persentase xilem aktif antara sampel yang ‘dipintal’ dan ‘dibilas’, kami akan menafsirkannya sebagai penyumbatan permanen; namun, jika ada perbedaan, kami akan menafsirkannya sebagai emboli.
Kami memperoleh data Palmer Severity Drought Index (PDSI) dan Climatic Water Deficit (CWD) dari TerraClim (Abatzoglu et al ., 2018 ; diakses masing-masing pada 21 Februari 2023 dan 24 Februari 2023) untuk lokasi aspen di sepanjang gradien iklim dari 01 Januari 2000 hingga 31 Desember 2022 (Gbr. S2 ). PDSI minimum dan CWD maksimum untuk tahun 2018, 2020, dan 2021 diekstraksi untuk lokasi setiap populasi aspen (Tabel S4 ). Kami memperoleh data normal 30 tahun untuk suhu dan presipitasi dari PRISM (PRISM Climate Group, Oregon State University, nd ), http://prism.oregonstate.edu , diakses 26 Oktober 2024) untuk periode Januari 1991 hingga September 2020. Kami menghitung jumlah hari di bawah titik beku dari setiap tahun dari suhu harian minimum PRISM dari 01 Januari 1991 hingga 31 Desember 2020 untuk mengeksplorasi potensi efek emboli beku-cair pada pola anatomi xilem.
Analisis statistik
Untuk semua analisis, kami menggunakan jumlah kuadrat tipe III dalam ANOVA untuk menguji perbedaan dalam kelompok cincin dan di seluruh populasi/lokasi dan melakukan perbandingan berpasangan dengan emmean (Lenth, 2024 ) kecuali dinyatakan lain. Jika efek interaksi tidak signifikan, efek tersebut dikeluarkan dari analisis dan ANOVA dijalankan ulang (Zuur et al ., 2009 ). Kecuali dinyatakan lain, data memenuhi asumsi model. Kami melakukan semua analisis statistik menggunakan perangkat lunak R untuk komputasi statistik versi R 4.2.2 (tim inti R, 2022 ).
Bahasa Indonesia : Untuk menguji apakah cincin berumur berbeda dan populasi dan lokasi berbeda berbeda dalam kemampuan mereka untuk menahan emboli, kami membandingkan nilai P50 dan PLC pada -1 dan -2 MPa. Kami melakukan ini untuk gradien iklim dan gradien elevasi. PLC pada -1 dan -2 dibandingkan karena ini adalah tekanan negatif tempat aspen biasanya beroperasi (LDL Anderegg et al ., 2013 ; Love et al ., 2019 ; Gambar S3 ). Untuk melakukan ini, kami memasangkan model linear dengan metrik PLC sebagai variabel respons dan populasi dan bin sebagai faktor. Untuk P50 , kami awalnya menghilangkan outlier menggunakan metode interkuartil, tetapi akhirnya memilih batas pada -4 MPa untuk outlier, karena ini adalah P50 minimum untuk aspen yang ditemukan di Love et al . ( 2019 ) untuk populasi aspen di wilayah yang sama. Untuk analisis sensitivitas menggunakan batas -3,5 MPa untuk outlier, lihat Gambar S4(a) . Kami melakukan transformasi logaritma data dan kemudian menyesuaikan model linier dengan metrik resistensi emboli sebagai variabel respons dan populasi serta bin cincin sebagai faktor untuk gradien iklim, dan elevasi sebagai faktor untuk gradien elevasi.
Untuk menguji apakah cincin-cincin yang berumur berbeda dan populasi-populasi dan lokasi-lokasi yang berbeda berbeda dalam kemampuan mereka untuk mengangkut air, kami membandingkan K s pada -1 dan -2 MPa. Untuk melakukan ini, kami memasangkan model linear dengan metrik konduktivitas sebagai variabel respons dan populasi dan bin sebagai faktor-faktor. Selain membandingkan K s , kami juga membandingkan persentase setiap cincin yang secara aktif berkontribusi pada pengangkutan air menggunakan perfusi zat warna. Kami memasangkan model linear dengan persentase xilem aktif sebagai variabel respons dan populasi dan bin sebagai faktor-faktor.
Kami kemudian melakukan serangkaian tes untuk memahami pola yang diamati dalam konteks gradien iklim. Pertama, kami menguji apakah perbedaan dalam K s dan PLC di antara tempat sampah dan populasi disebabkan oleh perbedaan diameter pembuluh. Untuk melakukan ini, kami menggunakan populasi dan tempat sampah sebagai faktor untuk memprediksi diameter pembuluh dengan ANOVA (perintah aov dalam R). Langkah yang sama ini diulang dengan diameter hidrolik ( D h ). Selanjutnya, kami menggunakan ANOVA tipe II untuk menentukan perbedaan dalam jumlah hari di bawah titik beku per tahun untuk setiap populasi. Kemudian, untuk menentukan apakah diameter pembuluh dan hari-hari di bawah titik beku berkorelasi, kami melakukan korelasi peringkat Spearman antara diameter pembuluh rata-rata dan jumlah rata-rata hari di bawah titik beku dalam populasi.
Untuk membandingkan persentase xilem aktif dalam keadaan aslinya, keadaan dipintal, dan keadaan terbuang, ANOVA tiga arah dilakukan di antara pita elevasi tempat cabang dikumpulkan, keadaan batang diwarnai (asli, terbuang, atau dipintal), dan wadah cincin.
Hasil
Gradien iklim elevasi
Kurva kerentanan spesifik cincin di antara ketinggian
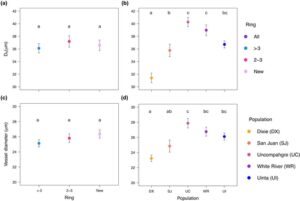
Kami tidak mendeteksi perbedaan signifikan dalam PLC pada -1 MPa di antara elevasi yang berbeda ( F 2,77 = 0,416, P = 0,6613). Namun, ada perbedaan signifikan dalam PLC di antara tempat penyimpanan ( F 3,77 = 5,027, P = 0,0031). Perbandingan berpasangan mengonfirmasi bahwa cincin yang lebih tua dari 3 tahun memiliki lebih banyak PLC pada -1 MPa daripada cincin terbaru dan luas penampang penuh (Gbr. 2a ). Pada -2 MPa, kami tidak menemukan perbedaan signifikan di antara elevasi dengan data dari semua tempat penyimpanan digabungkan ( F 2,78 = 2,842, P = 0,0643) atau di antara tempat penyimpanan cincin ( F 3,78 = 0,475, P = 0,7009; Gbr. 2a,b ), yang menunjukkan pembuluh darah yang mengalami emboli secara merata setelah -2 MPa. Kami tidak menemukan perbedaan signifikan dalam P 50 di antara kelompok cincin ( F 3,81 = 0,5562, P = 0,6455) atau elevasi ( F 2,82 = 0,5398, P = 0,5849; Gambar S5 ). Untuk hasil analisis tanpa kelompok yang memuat seluruh area penampang melintang, lihat Tabel S5 . Untuk kurva penurunan kerentanan dan konduktivitas dari gradien iklim elevasi, lihat Gambar S6 .
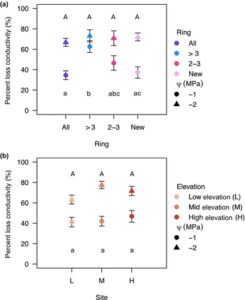
Perfusi pewarna di antara ketinggian
Kami tidak menemukan perbedaan signifikan dalam proporsi aktif xilem per bin di antara ketinggian (
= 0,415, df = 2, P = 0,813). Namun, terdapat perbedaan proporsi aktif xilem di setiap kelompok cincin (
= 43.888, df = 2, P < 0,001). Cincin yang paling baru memiliki proporsi xilem aktif yang lebih tinggi dibandingkan cincin yang berusia 2 dan 3 tahun dan cincin yang berusia lebih dari 3 tahun (Gbr. S7 ).
Gradien iklim antar populasi
Kurva kerentanan spesifik cincin di antara populasi
Kami membuat kurva kerentanan untuk setiap bin cincin untuk setiap populasi (Gbr. 3a,c ). Kami tidak menemukan perbedaan signifikan di antara populasi dalam P 50 saat menggabungkan semua bin ( F 4,103 = 1,2196, P = 0,3071). Namun, ada perbedaan di antara bin ( F 3,103 = 2,7094, P = 0,0490), dengan cincin terbaru memiliki P 50 yang jauh lebih negatif daripada cincin yang lebih tua dari 3 tahun ( P = 0,0443). Kami menemukan perbedaan signifikan di antara populasi dan bin pada -1 MPa. Cincin tertua (> 3 tahun) memiliki lebih banyak PLC pada -1 MPa daripada seluruh luas penampang, cincin berusia 2 hingga 3 tahun, dan cincin terbaru ( F 3,103 = 5,746, P = 0,0011; Gbr. 3b ). Ada perbedaan signifikan di antara populasi pada -1 MPa ( F 4,103 = 2.971, P = 0.0229; Gambar 3d ), karena San Juan memiliki PLC yang lebih tinggi daripada Uinta ( P = 0.0087). PLC pada -2 MPa berbeda di kedua tempat sampah ( F 3,110 = 2.734, P = 0.0471; Gambar 3b ) dan populasi ( F 4,110 = 3.230, P = 0.0151; Gambar 3d ). Cincin berusia dua dan tiga tahun memiliki PLC yang secara signifikan lebih tinggi pada -2 MPa daripada cincin terbaru. Baik San Juan maupun White River memiliki PLC yang secara signifikan lebih tinggi pada -2 MPa daripada Uinta. Untuk hasil analisis tanpa tempat sampah yang memuat seluruh area penampang, lihat Tabel S6 .
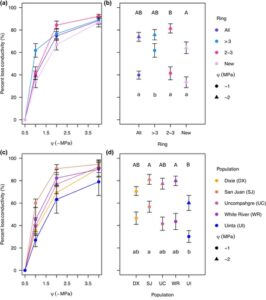
Kami juga membuat kurva penurunan konduktivitas untuk setiap bin cincin untuk setiap populasi (Gbr. 4a ,c ). Untuk Ks pada -1 MPa, kami menemukan bahwa bin tersebut berbeda secara signifikan satu sama lain ( F 3,112 = 40,5669, P < 0,001). Cincin terbaru memiliki Ks tertinggi pada -1, diikuti oleh semua cincin dan cincin berusia 2 hingga 3 tahun, dengan cincin tertua memiliki Ks terendah (Gbr. 4b ). Pada -1 MPa, beberapa populasi juga berbeda secara signifikan satu sama lain ( F 4,112 = 7,1981, P < 0,001), dengan Uinta memiliki Ks lebih dari dua kali lebih tinggi daripada San Juan dan Ks lebih dari empat kali lebih tinggi daripada Dixie (Gbr. 4d ) . Dixie memiliki Ks lebih rendah daripada semua populasi kecuali San Juan. Pada -2 MPa, bin-bin tersebut berbeda secara signifikan ( F 3,112 = 15,8692, P
< 0,001) dengan hanya cincin terbaru yang memiliki K s lebih tinggi daripada bin-bin lainnya (Gbr. 4b ). Populasi-populasi tersebut juga berbeda ( F 4,112 = 3,7278, P = 0,0069) dengan Uinta yang memiliki K s lebih dari 7 kali lebih tinggi daripada Dixie (Gbr. 4d ).
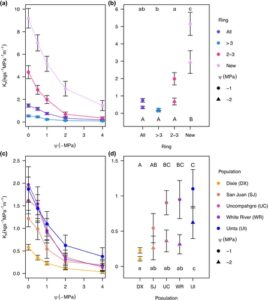
Untuk K max , kami mendeteksi perbedaan signifikan di antara bin ( F 3,112 = 147,426, P < 0,001; Gambar S8a ), di mana semua cincin berbeda secara signifikan satu sama lain. Cincin terbaru memiliki konduktivitas tertinggi per area, diikuti oleh cincin berusia 2 hingga 3 tahun, kemudian oleh semua cincin, dan cincin yang lebih tua dari 3 tahun memiliki konduktivitas terendah per area. Populasi juga berbeda secara signifikan satu sama lain ( F 4,112 = 21,650, P < 0,001; Gambar S8b ), dengan Uinta, White River, dan Uncompahgre memiliki konduktivitas tertinggi per area, San Juan di tengah, diikuti oleh Dixie yang memiliki K max terendah .
Diameter pembuluh darah
Kami tidak menemukan perbedaan signifikan dalam diameter pembuluh di antara cincin ( F 2,83 = 1,834, P = 0,166; Gambar 5c ). Namun, ada perbedaan signifikan dalam diameter pembuluh di antara populasi ( F 4,83 = 9,170, P < 0,001) dengan Uncompahgre memiliki diameter pembuluh yang secara signifikan lebih besar daripada Dixie dan San Juan. Semua populasi lainnya memiliki diameter pembuluh yang lebih besar daripada Dixie kecuali San Juan (Gambar 5d ). Untuk diameter hidrolik, juga tidak ada perbedaan signifikan di antara tempat sampah ( F 2,83 = 0,783, P = 0,461; Gambar 5a ), tetapi ada perbedaan di antara populasi ( F 4,83 = 18,371, P < 0,001). Semua populasi memiliki diameter hidrolik yang lebih besar daripada Dixie, diikuti oleh San Juan, dan tiga populasi lainnya tidak berbeda secara signifikan (Gambar 5b ).
Perfusi pewarna
Kami tidak mendeteksi perbedaan signifikan dalam proporsi xilem aktif di antara populasi (χ 2 = 5,456, df = 4, P = 0,244). Cincin berusia 2 hingga 3 tahun dan cincin terbaru tidak berbeda secara signifikan satu sama lain, tetapi cincin yang berusia lebih dari 3 tahun memiliki proporsi xilem aktif yang secara signifikan lebih rendah (χ 2 = 38,463, df = 2, P < 0,001; Gambar S9 ).
Hari-hari beku di antara populasi dan korelasi dengan diameter pembuluh darah
Terdapat perbedaan signifikan dalam jumlah hari di bawah titik beku per tahun di antara populasi ( F 4,145 = 39,137, P < 0,001). Hutan Nasional Dixie dan Uinta memiliki hari beku yang jauh lebih banyak per tahun daripada Hutan Nasional San Juan, Uncompahgre, dan White River. Hutan Nasional San Juan dan White River memiliki jumlah hari di bawah titik beku tertinggi berikutnya, dan Uncompahgre memiliki jumlah hari di bawah titik beku paling sedikit (Gbr. S10 ). Terdapat korelasi signifikan antara jumlah rata-rata hari beku per tahun dan diameter pembuluh rata-rata, karena diameter pembuluh menurun dengan jumlah hari beku yang lebih banyak per tahun ( S = 180 526,
= −0,486, P < 0,001; Gambar S11 ).
Validasi metode tambahan
Untuk pengujian yang melihat seberapa banyak zat warna bergerak melintasi batas cincin, kami menemukan bahwa rata-rata 1,95% dari area xilem yang direkatkan memiliki zat warna yang bocor melintasi cincin (Gbr. S12 ). Untuk pengujian zat warna yang membandingkan berbagai keadaan konduktivitas (dibilas, asli, atau dipintal), tidak ada interaksi di antara variabel independen. Situs rendah dan tinggi berbeda secara signifikan satu sama lain ( F 1,105 = 12,333; P < 0,001), dengan situs tertinggi memiliki xilem yang lebih aktif daripada situs yang lebih rendah ( P < 0,001). Keadaan batang yang mengalami perfusi zat warna (dibilas, asli, dipintal pada −2 MPa) berbeda secara signifikan ( F 2,105 = 3,211, P = 0,044), di mana batang yang dibilas memiliki area xilem yang lebih aktif secara signifikan daripada yang dipintal pada −2 MPa. Namun, tidak ada perbedaan antara ‘flushed’ dan ‘native’, juga tidak ada perbedaan antara ‘native’ dan batang yang dipintal pada -2 MPa. Ini menunjukkan bahwa xilem yang tidak aktif dalam keadaan asli tanaman kemungkinan besar disebabkan oleh penyumbatan permanen, tetapi pada tekanan negatif, emboli adalah penyebab xilem yang tidak aktif. Kelompok cincin berbeda secara signifikan satu sama lain ( F 3,105 = 66,631, P < 0,001), dengan cincin terbaru memiliki persentase area xilem yang diwarnai secara signifikan lebih banyak, diikuti oleh cincin berusia 2 dan 3 tahun, diikuti oleh seluruh area penampang melintang dan kemudian diikuti oleh cincin tertua (semua nilai P < 0,001). Diskusi Kekeringan yang lebih sering terjadi karena perubahan iklim menyoroti kebutuhan mendesak untuk lebih memahami bagaimana beberapa kekeringan dan penuaan xilem memengaruhi transportasi air tanaman. Sebagian besar kurva kerentanan dibuat menggunakan seluruh luas penampang sampel; dengan demikian, kurva ini tidak memperhitungkan perbedaan di antara cincin pertumbuhan tahunan dan mungkin melebih-lebihkan kontribusi cincin tua terhadap konduktansi batang secara keseluruhan (Melcher et al ., 2003 ; Fukuda et al ., 2015 ). Dalam studi ini, kami mengembangkan metode baru untuk mengukur fungsi xilem spesifik cincin dan menemukan bahwa cincin dengan usia yang berbeda berbeda dalam kemampuannya untuk menahan emboli dan mengalirkan air. Dengan menggunakan data dari lima populasi Populus tremuloides yang tumbuh dalam rezim iklim yang kontras dan tiga populasi melintasi gradien elevasi/curah hujan, kami menemukan dukungan untuk dua hipotesis pertama kami bahwa cincin yang lebih tua tidak hanya mengalami konduktivitas hidrolik spesifik area yang lebih rendah tetapi juga peningkatan kerentanan terhadap emboli, yang menunjukkan pemisahan trade-off keamanan vs efisiensi di seluruh cincin individual. Kami juga menemukan dukungan untuk hipotesis ketiga kami bahwa khususnya di lokasi dan populasi yang lebih kering, kerentanannya lebih tinggi dan konduktivitasnya lebih rendah. Secara keseluruhan, hasil ini menunjukkan bahwa xilem yang mengalami stres kekeringan terus-menerus tampaknya mengalami kerusakan dan bahwa xilem terbaru yang belum mengalami stres kekeringan terus-menerus memberikan kontribusi paling besar terhadap transportasi air dan ketahanan terhadap emboli. Kontribusi spesifik cincin terhadap toleransi kekeringan dan transportasi air Kami menemukan bahwa cincin terbaru memiliki P 50 yang lebih tahan daripada cincin yang lebih tua dari 3 tahun, dan bahwa kumpulan cincin mencapai tingkat PLC yang berbeda pada -1 dan -2 MPa. Cenderung ada perbedaan yang lebih besar di antara cincin dalam rentang tegangan xilem ini (Gbr. 3a ), yang penting karena ini adalah rentang potensial air tengah hari musim panas yang sangat khas untuk aspen di SJNF (Anderegg et al ., 2013b , LDL Anderegg et al ., 2013 , Anderegg & Hillerislambers, 2016 , Love et al ., 2019 ; Gbr. S3 ; untuk PLC asli, lihat Gbr. S13 ). Hasil kami mengungkapkan bahwa cincin berusia 1-3 tahun paling tahan terhadap emboli pada potensial air ini dan bahwa cincin yang lebih tua dari 3 tahun tidak dapat beroperasi dengan aman dalam kondisi musim panas rata-rata (PLC di atas P 50 pada potensial air rata-rata). Hasil ini konsisten dengan Pratt et al . ( 2020 ) yang menemukan bahwa batang berumur 1, 2, dan 3 tahun memiliki kesesuaian yang baik dalam kurva dehidrasi benchtop. Kemungkinan membran pit dari pembuluh yang lebih tua melemah karena kelelahan kavitasi, sehingga penyebaran emboli lebih mudah melalui ‘lubang bocor’ (Sperry et al ., 1991 ; Fukuda et al ., 2015 ; Hillabrand et al ., 2016 ). Selain itu, ada kemungkinan juga bahwa cincin yang lebih tua ini tersumbat karena pembentukan tyloses. Namun, pembentukan tyloses sering dikaitkan dengan embolisme, kerusakan xilem (Sperry et al ., 1991 ; Taylor et al ., 2002 ), respons luka (Leśniewska et al ., 2017 ), dan kami percaya bahwa stres kekeringan dan embolisme dari paparan kekeringan sebelumnya berperan dalam proses ini. Pada saat seluruh luas penampang mencapai P 50 , cincin tertua rata-rata mengalami embolisasi sebesar 67%. Dibandingkan dengan kayu keras lainnya, pencitraan resonansi magnetik menunjukkan bahwa pada saat seluruh batang mencapai kehilangan konduktivitas sebesar 50%, cincin yang lebih tua dari tahun berjalan sudah mengalami embolisasi sebesar 100% (Fukuda et al ., 2015) .). Kami secara umum menemukan bahwa kurva untuk seluruh luas penampang tampak merupakan rata-rata antara kurva cincin terbaru dan cincin yang berusia lebih dari 3 tahun dan bahwa seluruh luas penampang tidak berbeda dalam PLC dari cincin terbaru pada -1 dan -2 MPa. Hal ini karena cincin terbaru memberikan kontribusi paling besar terhadap konduktivitas dan dengan demikian mendominasi sinyal pengukuran. Temuan ini memiliki implikasi metodologis yang penting, karena dalam beberapa kasus, mungkin tidak perlu melakukan proses padat karya untuk melakukan kurva kerentanan khusus cincin jika PLC dari cincin terbaru diinginkan, setidaknya pada spesies ini atau spesies serupa. Cincin tua (> 3 thn) tidak banyak berkontribusi pada konduktivitas seluruh batang, dan cara pengukuran Ks biasanya, sebagai area yang dinormalisasi oleh seluruh batang, kemungkinan merupakan perkiraan yang lebih rendah dari Ks area xilem aktif karena penyertaan xilem yang tidak aktif dalam pengukuran. Hipotesis kami bahwa cincin terbaru akan menghantarkan lebih banyak air dan bahwa cincin lama akan menghantarkan lebih sedikit air didukung oleh area xilem yang diwarnai dan Ks . Cincin baru, terutama sepertiga pertama xilem dari kambium, lebih efisien dalam transportasi air dan berkontribusi pada setengah dari transportasi air batang pada spesies peluruh (Melcher et al ., 2003 ; Dang et al ., 2014 ; Fukuda et al ., 2015 ). Ini mungkin karena cincin yang lebih baru bertanggung jawab untuk transportasi air ke daun (Melcher et al ., 2003 ; Fukuda et al ., 2015 ), sementara cincin yang lebih tua dari 3 tahun mengandung xilem yang jarang digunakan untuk transportasi air dan malah lebih sering digunakan untuk penyimpanan air (Tyree & Zimmermann, 2002 ). Namun, pola-pola ini sering kali khusus untuk klade atau spesies, dan dalam kasus aspen, daun hanya disuplai air oleh cincin terbarunya, dan kemungkinan ada resistensi yang sangat tinggi untuk pergerakan air melintasi cincin, tetapi penelitian lain telah menemukan jalur melintasi cincin pertumbuhan (Pratt et al ., 2020 ). Kami menemukan bahwa beberapa makalah lain yang menggunakan berbagai metode di banyak spesies menemukan tren serupa bahwa hubungan radial melintasi batas cincin minimal (Domec et al ., 2005 ; Barnard et al ., 2013 ; Wason et al ., 2019 ; Petit et al ., 2023 ).
Meskipun metode pembuatan kurva kerentanan ini secara umum berhasil memisahkan perbedaan konduktivitas dan kerentanan di antara lingkaran pertumbuhan tahunan, beberapa peringatan perlu diperhatikan. Salah satunya adalah bahwa kurva kerentanan tidak dibuat dengan titik pengukuran potensial air beresolusi tinggi seperti yang biasanya disukai. Tempat sampah potensial air beresolusi rendah disebabkan oleh kebutuhan untuk memotong lem sebelum setiap putaran di sentrifus. Memotong batang terlalu sering akan membuat batang terlalu pendek untuk rotor, jadi kurva-V disingkat untuk menyertakan lebih sedikit putaran. Selain itu, kami menghitung konduktivitas untuk cincin berusia 2 hingga 3 tahun melalui pengurangan konduktivitas cincin terbaru dari konduktivitas tiga cincin terbaru. Cara ini mungkin tidak seakurat mengukur konduktivitas tempat sampah ini secara langsung, tetapi dilakukan untuk mencegah lebih banyak perekatan dan pemotongan.
Variasi melintasi gradien lingkungan yang luas
Bahasa Indonesia : Saat membandingkan antar populasi dan melintasi gradien elevasi, kami tidak menemukan perbedaan signifikan dalam P50 , tetapi ada perbedaan antar populasi pada -1 dan -2 MPa. San Juan memiliki beberapa xilem paling rentan pada -1 dan -2 MPa, sementara Uinta memiliki beberapa xilem paling resistan pada tekanan ini. Hasil ini serupa dengan yang ditemukan dalam Kerr et al . ( 2023 ) di mana populasi yang lebih kering lebih rentan terhadap emboli daripada populasi yang lebih basah. Ini mungkin menunjukkan bahwa populasi kering (Dixie dan San Juan NF) telah mengalami kelelahan kavitasi (Hacke et al ., 2001 ). Penelitian ekstensif telah mendokumentasikan mortalitas aspen yang disebabkan oleh kekeringan yang substansial di wilayah ini dan mendukung gagasan bahwa populasi ini mengalami penurunan hidrolik (Huang & Anderegg, 2012 ; Anderegg et al ., 2013a , b ).
Mengenai konduktivitas, Dixie memiliki konduktivitas terendah pada -1 MPa, diikuti oleh San Juans, dengan tiga populasi lainnya tidak berbeda dalam konduktivitas. Perbedaan konduktivitas di antara populasi dapat dijelaskan sebagian oleh ukuran diameter pembuluh, karena pembuluh yang lebih besar cenderung lebih konduktif secara hidrolik (Sperry et al ., 2006 ). Langkah masa depan yang menarik akan mencakup memisahkan apakah perbedaan dalam K h dimediasi oleh diameter pembuluh atau melalui kuantitas xilem aktif pada batang yang sama. Selain itu, kami mencatat korelasi antara ukuran diameter pembuluh dan jumlah hari di bawah titik beku (Gbr. S10 ), yang telah diamati pada spesies lain (Davis et al ., 1999 ). Baik CWD dan tingginya jumlah hari di bawah titik beku menunjukkan bahwa Hutan Nasional Dixie dapat mengalami baik stres kekeringan maupun stres beku-cair (Ewers et al ., 2003 ), yang dapat berinteraksi untuk menciptakan stres xilem tambahan (Langan et al ., 1997 ; Feng et al ., 2015 ). Sementara konduktivitas hidrolik dan diameter pembuluh dianggap berkorelasi dengan ketersediaan air, diameter pembuluh malah dibatasi oleh stres beku-cair dan lokasi yang lebih menengah lebih seimbang antara stres kekeringan dan stres beku-cair.
Kesimpulan
Sebagai kesimpulan, temuan kami mengungkap bahwa ada perbedaan substansial dalam cincin dengan berbagai usia untuk mengalirkan air dan menahan emboli. Secara umum, cincin terbaru adalah yang paling tahan pada potensi air musim panas standar dan berkontribusi paling besar terhadap konduktansi air, sementara cincin yang lebih tua dari 3 tahun sangat rentan dan tidak banyak berkontribusi terhadap konduktansi. Pola-pola ini terlihat pada skala luas dalam gradien elevasi dan gradien iklim. Di gradien iklim, populasi yang mengalami stres kekeringan paling banyak lebih rentan, yang menyoroti akumulasi sinyal kerusakan daripada sinyal aklimatisasi. Selain itu, interaksi antara stres kekeringan dan pembekuan-pencairan mungkin berdampak pada konduktivitas dan ukuran diameter pembuluh. Pemahaman yang lebih luas tentang bagaimana perubahan dalam transportasi air khusus cincin memengaruhi fluks air dan kelangsungan hidup seluruh tanaman, serta perubahan konduktivitas di seluruh cincin dalam taksa yang berbeda, sangat dibutuhkan untuk menjelaskan bagaimana lebih banyak iklim ekstrem akan memengaruhi hutan dalam beberapa dekade mendatang.


Leave a Reply